
William LEVENGOOD,
et al
Electroculture
Electroculture
http://www.iccra.org/levengood/allotherpub.htm

William LEVENGOOD
W.C. LEVENGOOD, JUDITH BONDIE and CHI-LING CHEN,
Biophysical Research Department, Sensors, Inc.
3908 Varsity Drive, Ann Arbor, Michigan 48104
Seed Selection for Potential
Viability.
A simple method is presented for selecting individual seeds for growth and vigour prior to germination. The selection is based on the measurement of an electric current originating during the initial stage of seed imbibition. After selecting, seeds may be returned to the quiescent state without affecting viability. Results of both laboratory germination and field experiments demonstrate that high germination rates, more extensive growth, higher yields, and fewer defective plants are associated with small currents. Seeds of low viability and high currents may be removed to upgrade the germination and vigour. Results are presented from several varieties of Phaseolus vulgaris and from three other plant species. Experiments are discussed which indicate that seeds graded by current levels possess markedly different respiration rates.
Method and apparatus for enhancing
growth characteristics of seeds using ion-electron
avalanches
US6023880
[ PDF ]
US6023880
[ PDF ]
Also published as: US5740627 // WO9811770 // JP2000502911 // JP3213329
Abstract
A method and apparatus for treating seeds with self-organized avalanches of electrons between electrodes (11, 12) as a cathode and an anode with seeds (13) between the anode and cathode or on the anode. Apparatus circuit (200) in a box (20) provides simultaneous DC and AC between the electrodes which creates the avalanche of electrons which project into the seeds. The seeds must be stored before planting. The seeds so treated have enhanced growth characteristics.
BACKGROUND OF THE INVENTION
(1) Field of the Invention
The present invention relates to a method and apparatus for treating seeds, thereby reproducibly enhancing rate and uniformity of seed germination, early growth, root growth, maturity, and yield in food crops and other plants. These results are achieved by exposing seeds or growing plants to uniform, spontaneously-organized pulses of ion-electron avalanches. One important aspect is allowing a period of several weeks storage before planting thereby allowing internal, biochemical changes to take place at the cellular level within the seed. The present invention also relates to a quality-control method and apparatus for selecting optimal treatment parameters with the avalanches of ions and electrons for each variety of seed.
(2) Description of Related Art
Almost since the discovery of the commercial use of electricity, experimenters have tried to electrically influence plant growth. Various prior art experimenters have claimed positive results from exposing growing plants to electrical stimulation in situ. A wiring network over a field of growing crops is not cost-effective or practical on a commercial scale, and such techniques have not been adopted by farmers.
Some prior art experimenters have attempted to avoid the prohibitive cost of wiring a field by applying electromagnetic treatments to seeds before planting. Despite reports of increased growth and, in some cases, increased yield, these results have proven difficult to repeat and have not achieved commercial use. Parry (U.S. Pat. No. 2,308,204 (1943)) describes the use of an oscillating DC voltage to treat seeds to increase germination of the seeds. There is no indication of improved plants. Jonas (U.S. Pat. No. 2,712,713 (1955)) and others exposed seeds to high frequency oscillating fields between 30 MHz and microwave range, claiming faster and more uniform germination. Jonas stated that the work of others along similar lines have been impossible to repeat and confirm. The patent describes only increased germination of the seeds. Amburn (U.S. Pat. Nos. 3,675,367 (1972) and 3,765,125 (1975)) exposed seeds to magnetic fields, claiming increased germination rate as an effect. Because of unreliability and non-reproducibility, none of these methods have achieved widespread commercial acceptance.
Levengood (U.S. Pat. No. 3,822,505 (1974)) describes an apparatus for genetically altering plant cells using combined electrical and magnetic fields. The electrical field is static. There was alteration in the growth of seeds, but the method was not repetitively effective from batch to batch of seeds. Another patent to Levengood (U.S. Pat. No. 3,852,914 (1974)) describes a method for testing seeds for viability, by measuring pregermination tissue conductivity.
Schiller et al (U.S. Pat. No. 4,633,611 (1987)) describe treating seeds to disinfect them with low energy electrons using an electron gun. The radiation dosages are quite high and the acceleration voltages are between 25 and 75 kV. The use of high energy ionizing radiation can cause damage to chromosomes and resultant genetic change which poses complications for use in open fields. There is no indication that the growth of the plant is enhanced on a reproducible basis. Yoshida (U.S. Pat. No. 4,758,318 (1988)) describes using a pulsating direct current to prevent mold. The voltages were 300 to 20,000 V DC which were pulsed. This method is not practical on a large scale and the results were variable. Liboff et al (U.S. Pat. No. 5,077,934 (1992)) describe the use of magnetic fields with plants in the soil. This method is not practical.
Levengood (U.S. Pat. No. 5,288,626 (1994)) describes genetically transferring DNA between plants using a constant DC voltage. This is also described in Bioelectrochemistry and Bioenergetics (1991). These are techniques for producing genetically altered plants.
Other patents of general interest are Saruwatari (U.S. Pat. No. 4,188,751 (1980)) relating to magnetic treatment; Weinberqer (U.S. Pat. No. 3,703,051 (1972)) relating to ultrasound; U.S. Pat. No. 3,940,885 (1976) relating to microwaves.
One system which used an A.C. ripple in a D.C. current to produce pulses is Tellefson (U.S. Pat. No. 5,117,579 (1992)). Pulses of ions were produced from wire brush emitters to flood growing plants in a field. The method is not used with seeds.
There is clearly a need for a reproducible and reliable method for treating seeds to enhance their growth characteristics. The prior art methods have not met this need since no such method is used commercially.
OBJECTS
It is therefore an object of the present invention to provide an improved, reproducible method and apparatus for enhancing the growth characteristics of seeds. It is further an object of the present invention to provide such a method which is simple, reliable and economical to perform. Further still it is an object of the present invention to provide a method and apparatus for detecting whether or not the treated seeds have been effectively improved in their growth characteristics by the method and apparatus for enhancing growth characteristics Further still, it is an object of the present invention to provide a method and apparatus which allows monitoring during treatment of the effectiveness of the apparatus for performing the treatment. These and other objects will become increasingly apparent by reference to the following specification and the drawings.
BRIEF DESCRIPTION OF DRAWINGS
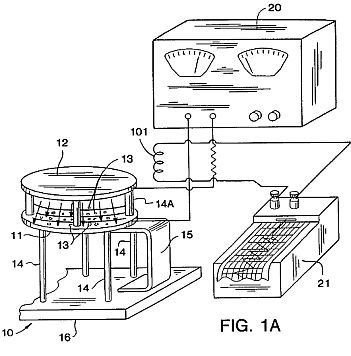
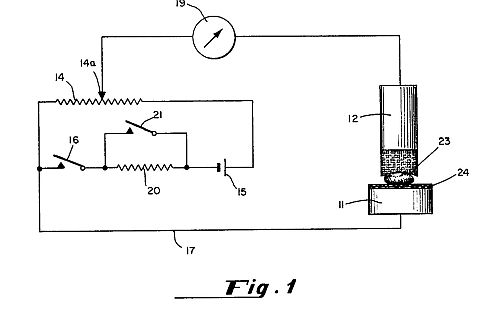
FIG. 1A is a schematic view of the apparatus of the present invention for producing controlled, spontaneous, electrostatic pulses which form the organized electron avalanches between an anode electrode 11 supporting seeds 13 and a cathode electrode 12.
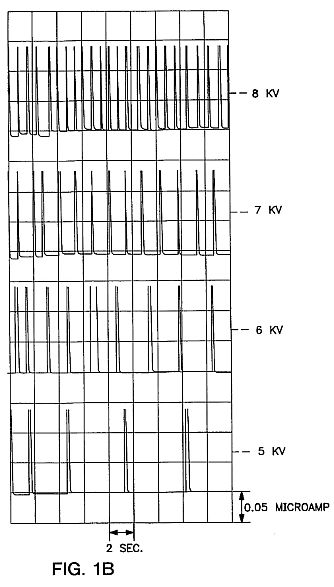
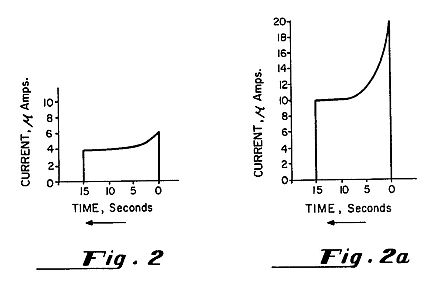
FIG. 1B is a charted graph showing organized electron avalanches produced in the apparatus of FIG. 1A with different DC voltages (relative humidity 26%; p =1009.3 mb).
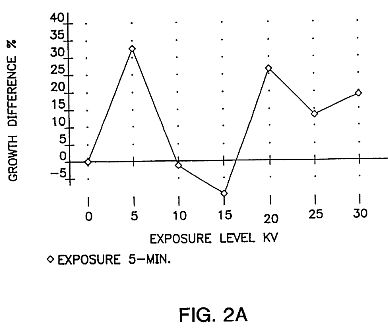
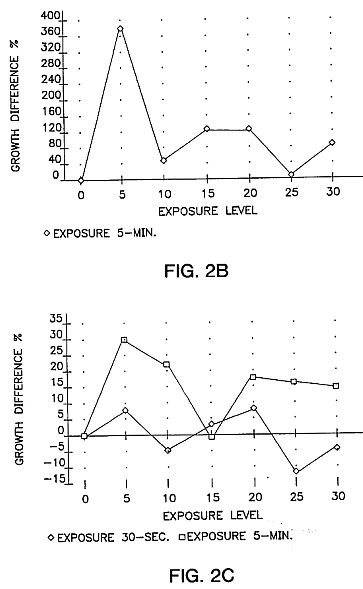
FIGS. 2A, 2B and 2C are graphs showing growth differences in tomatoes, pepper and carrot using a DC voltage for five minutes in the apparatus of FIG. 1A with seeds stored for 35 or 36 days. Germination data was taken at the 12-day growth stage and represents hypocotyl extension (seedlings placed under grow lights at 4-day development). The data was compared with two control sets in each test series. FIG. 2A shows tomato seeds tested 35 days after exposure. FIG. 2B shows pepper seeds tested 35 days after exposure. FIG. 2C shows carrot seeds tested 36 days after exposure. As can be seen, similar curve shapes appear in the 5-minute exposure data. In every case the maximum peak is at the 5-kV level, with a secondary peak at 20-KV.
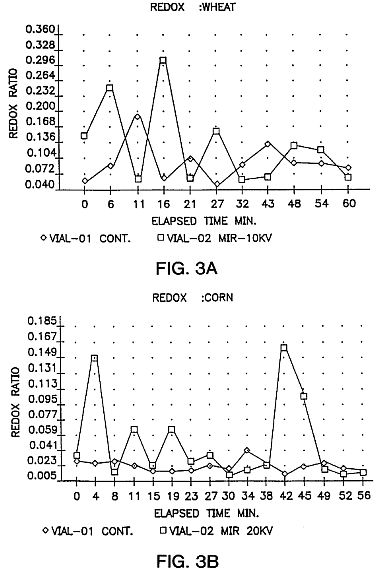
FIGS. 3A and 3B are graphs showing redox ratio (ratio of active anions to cations) changes in developing wheat and maize seedlings over a 60 minute test interval in both untreated, control seed and in seed exposed to the spontaneously organized ion-electron avalanches, with avalanche exposure of 30 seconds at 10 kV (FIG. 3A) and 20 kv (FIG. 3B) The seeds were stored for eight (8) days. The leaf tissue between electrodes 11 and 12 was tested after 12 days under a grow light.
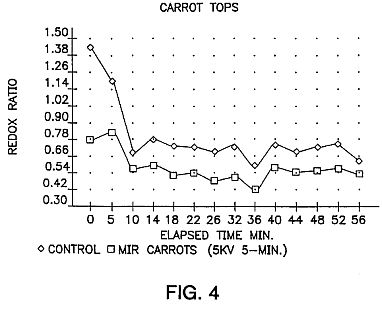
FIG. 4 is a graph showing redox ratio changes in mature, field grown carrot foliage from both untreated control seeds and seeds exposed to ion-electron avalanches at 5 kV for 5 minutes and stored for 81 days before planting. Redox Ratio: FIG. 4 shows redox ratios of MIR-treated carrots to be lower than that of untreated controls, when measured after the plants develop to the mature autotrophic phase. The redox potential is determined from exudate from the seeds.
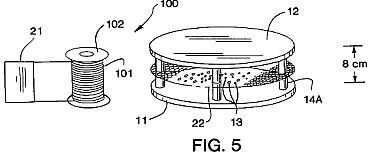
FIG. 5 is a schematic view of an apparatus 100 with a probe coil 101 for examining the induced-energy wave form from the ion-electron avalanche pulses produced by the apparatus of FIG. 1A. The coil 101 had 80,000 turns of #40 copper wire and was approximately 8 cm in diameter and 10 cm long on core 102.
The upper part of FIG. 6 is a graph showing the induced magnetic field in the coil 101 of FIG. 5 produced by the electron avalanches shown in the lower portion of FIG. 6. This gives a direct reading of the current between the electrodes 11 and 12 of FIG. 1A at an applied potential of 5 kV.
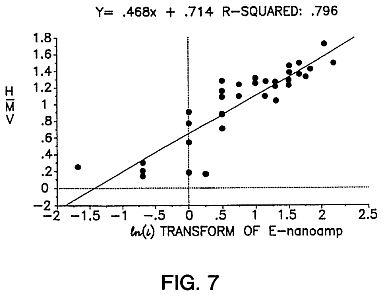
FIG. 7 is a graph showing an exponential correlation between the electron pulsed current between electrodes 11 and 12 and the magnetic field potential induced in the coil 101.
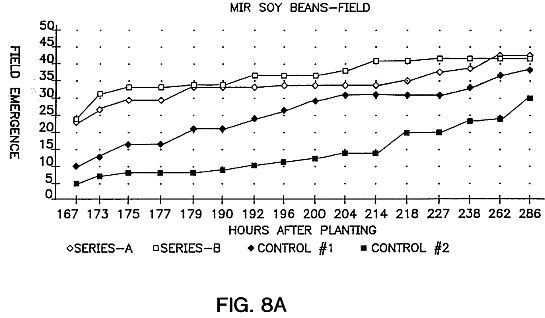
FIG. 8A is a graph showing 1995 field emergence rates in avalanche-exposed soybeans versus two control series. The seeds were Var. PS-202 (total of 48 seeds per test series). Series A: 5 kV, 5 min. Series B: 10 kV, 5 min. The seeds were stored for 86 days after treatment before planting.
FIG. 8B and 8C are graphs showing 1995 field emergence rates in two varieties of avalanche exposed sweet corn seed versus their controls. The seeds were stored for 56 days after treatment before planting.
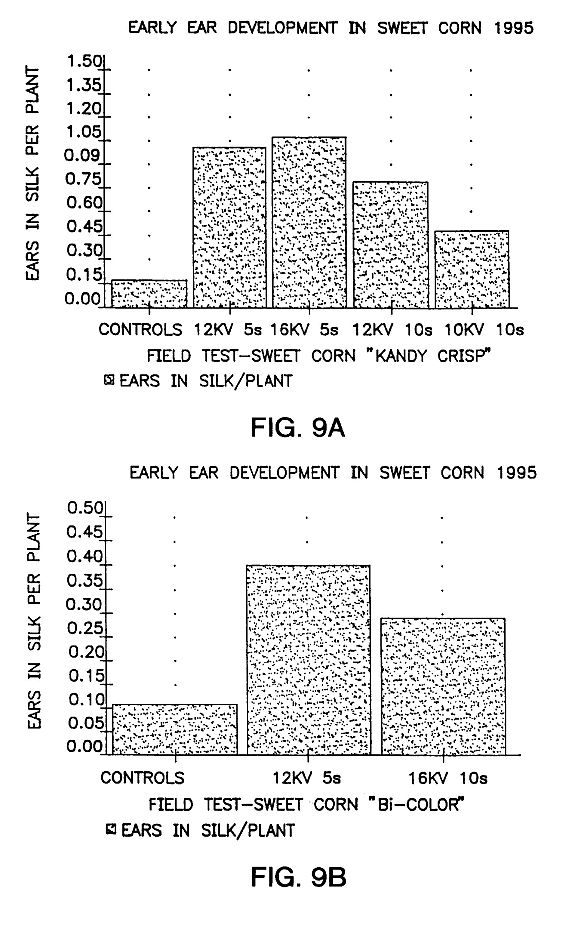
FIGS. 9A and 9B are graphs showing fruit or ear development in two varieties of 1995 field-grown sweet corn versus their controls. The seeds were stored for 56 days after treatment.
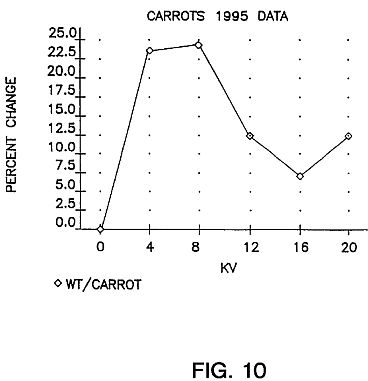
FIG. 10 is a graph showing carrot foliage yields in 1995 as a function of avalanche-inducing voltages. The field plot data is based on percent change in fruit relative to controls. Each point is a mean of a series of seeds exposed at 10 sec., 30 sec., 5 min. and 30 min. at the kv level indicated. The seeds were stored for 81 days before planting.
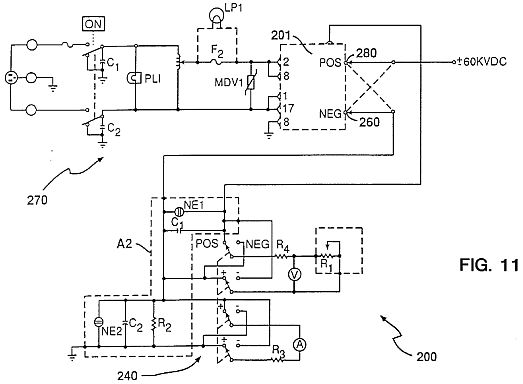
FIG. 11 is a circuit diagram 200 in box 20 of apparatus 10 for producing the spontaneous organized electron-ion avalanche pulses.
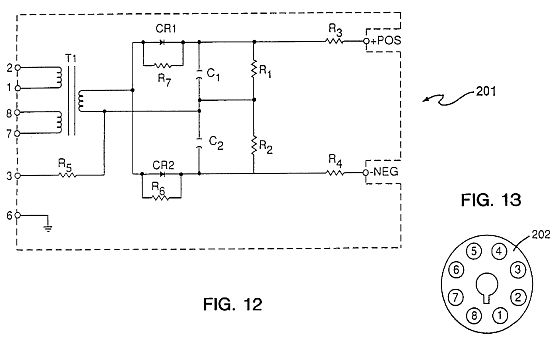
FIG. 12 is a circuit diagram for a power pack nodule 201 as shown in FIG. 11 in circuit 200 with the organized electron avalanches used in the method of the present invention.
FIG. 13 is a connector for the power pack nodule 201 of FIGS. 11 and 12.
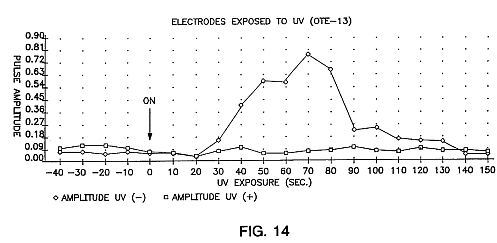
FIG. 14 is a graph showing changes in avalanche pulse amplitude as a result of photon-released electrons generated by ultraviolet light exposure at the cathode. There is no effect from exposing the anode, as we would expect from theoretical considerations.
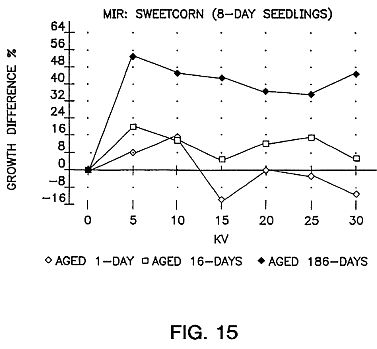
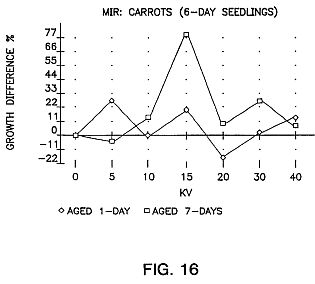
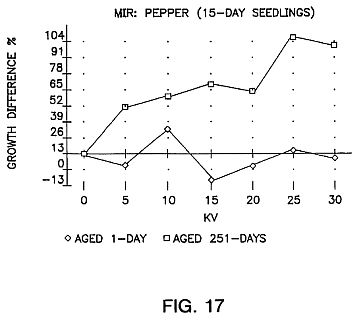
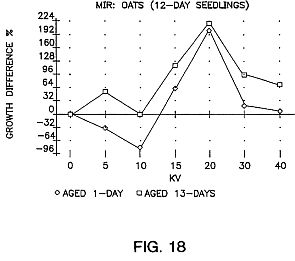
FIGS. 15, 16 and 17 are graphs showing the results of aging of the seeds for sweet corn (G18-86), carrots, pepper and oats with an exposure time of 25 seconds.
FIG. 18 is a graph showing the results of treating seeds in the panicle.

DESCRIPTION OF PREFERRED EMBODIMENTS
The present invention relates to a method for treating a seed to enhance growth characteristics of the seed which comprises: providing the seed between a pair of spread apart electrodes as an anode and a cathode having a gap between them and with seed on or adjacent to the anode; applying a direct current (DC) voltage to the anode and the cathode using a power supply with an output voltage with an impressed alternating current AC ripple on the output voltage so as to produce self-organized, or pulsed avalanches of electrons moving from the cathode towards and into the seed between the anode and cathode or on the anode for a period of time which enhances the growth characteristics of the seed; and storing the seed for a period of time before planting sufficient to allow the seed to provide the seed with the enhanced growth characteristics.
The present invention also relates to a seed produced by providing a space between an anode with the seed and the cathode, exposing the seed to pulsed avalanches of electrons produced by applying a DC voltage, with an AC ripple impressed upon the DC voltage, to spaced apart electrodes using a power supply with an impressed AC ripple in the output voltage to produce self-organized pulsed avalanches of ion-electrons which move towards and into the seed, and then storing the seed before planting.
The present invention also relates to a plant produced from a seed produced by exposing the seed to pulsed avalanches of electrons produced by providing spaced apart electrodes which are an anode and a cathode with the seed between the anode and cathode or on the anode, applying a DC voltage with impressed AC ripple to the spaced apart electrodes to produce self-organized avalanches of electrons which move towards the anode and into the seed before planting.
The present invention also relates to an apparatus for detecting the presence of pulsed avalanches of electrons in an apparatus for treatment of a seed which comprises: a solenoid coil with multiple turns which is adapted to be positioned adjacent to a pair of spaced apart electrodes including an anode supporting the seed; and detection means for detecting an induced current in the coil.
The present invention relates to an apparatus for treating a seed to enhance the growth characteristics of the seed which comprises: a pair of spaced apart electrodes as an anode and as a cathode having a gap between them wherein the seed is to be supported on or adjacent to the anode; voltage generating means for simultaneously supplying a direct current (DC) voltage to the anode and the cathode using a power supply with an output voltage with an impressed alternating current AC ripple as the output voltage so as to produce organized, pulsed avalanches of electrons moving from the cathode towards and into the seed on the anode for a period of time which enhances the growth characteristics of the seed; and coil means with multiple turns mounted adjacent to the spaced apart electrodes which detects pulsed avalanches of electrons; and recording means for recording the pulsed avalanches of electrons as detected by the coil means.
The present invention relates to a method for significantly improving the rate and uniformity of germination and early growth, as well as increased yield, in plants, particularly commercial crops, by a cost-effective treatment of the seeds using electron avalanches in a manner that can be reliably duplicated, and lends itself well to commercial exploitation. The method provides an apparatus for exposing seeds to organized avalanches of electrons from a flat electrode.
The seeds 13 are placed directly on top of a horizontal, flat aluminum (or other metal) plate or electrode 11 which is an anode spaced from an electrode 12 which is a cathode so that the electrode 11 is the bottom most of the two parallel electrodes 11 and 12. Alternatively, the seeds can be placed on a non-conducting screen 22 (FIG. 5) elevating them above the anode electrode 11. For all results listed here, the elelectrodes 11 and 12 used were round and 30 cm in diameter. Other shapes and sizes of electrodes can be used, though this may change the effective voltage levels. The electrodes 11 and 12 are supported by legs 14 and 14A made of a dielectric material. The bottom electrode 11 can take a variety of forms, such as a metal conveyor belt (not shown).
A high-voltage DC power supply 20 providing positive current is connected to the bottom electrode (anode) 11, while the top electrode 12 (cathode) is grounded. Improved results are obtained if the DC power supply contains an organized 60 or 220 Hertz ripple in the DC. Other than such an AC trace and its resultant ripple, there is no other oscillation of the DC current. This distinguishes the apparatus from prior art systems which use a voltage oscillator, usually in the megahertz range or higher.
Due to conductivity of the air between the electrodes 11 and 12, organized avalanches of electrons travel from the negative electrode 12 (cathode) to the positive electrode 11 (anode). These electron avalanches register as pulses on the monitoring equipment described hereinafter. When a "clean" signal DC power supply is used, both the frequency and amplitudes of the ion-electron avalanches are lower and more irregular. When a power supply with AC ripple is used, the avalanches form in regular self-organized, discrete pulses. These avalanche pulses commonly occur in the 0.1 to 30 Hz range between the electrodes 11 and 12 and are a product of voltage gradient and conductivity of the air between the electrodes 11 and 12, not of an artificial oscillator. The term "self-organized" means that there is a discharge between the electrodes 11 and 12 dependent upon the voltage and the environmental conditions between the electrodes 11 and 12.
The best results have been obtained when the electrodes 11 and 12 are supported on dielectric legs 14 on a plastic-topped table 16 and the bottom electrode 11 is grounded to the tabletop by a feedback loop 15 of a conductive metal. When the feedback loop 15 is added, the same electrode system produces pulses of very similar frequency to those obtained without the loop, but of significantly increased amplitude. The reason for this is that the table top 16 appears to function as a feed-back loop type of capacitor.
It has been found that an avalanche inducing voltage improving the seeds of some plant varieties was ineffective or actually harmful to seeds of other varieties. Likewise, the duration of the seed's exposure to the electron avalanches is important and variable. The diagnostic process to select the best times and voltages is also important. Finally, the waiting period before planting, and considerations of moisture in the air and seed temperature are important. The present method works well on seeds dried to normal levels for commercial storage and at temperatures above 40 DEG C. Monitoring apparatus, described later, can be used to adjust for altered air conductivity due to changes in relative humidity.
The method of the present invention is referred to as Molecular Impulse Response, or MIR. A specific type of impulse from an electron produces a molecular response in the seed which ultimately results in significantly-improved seed performance, when it is applied in the following manner, including but not limited to:
A.) Electrodes and Power Supply: Using a spacing between electrodes 11 and 12 (preferably 8 cm although other spacings, preferably between about 1 and 20 cm, can be used but will alter the effective voltages) and inducing a voltage gradient between the electrodes of about 2 kV or more (other voltages can be used up to, but below, the electrical breakdown voltage in air corona discharge) results in the production of organized electron avalanches which take the form of sharp, regular electrical conductivity pulses of relatively uniform amplitude in the air between the electrodes 11 and 12 (as traced on a chart recorder system 21 as shown in FIG. 1A) Such spontaneously organized electron avalanches have been described in the scientific literature, most notably by Nasser, as examples of a low density, low energy plasma in air at ambient pressure. (Source: E. Nasser, "Fundamentals of Gaseous Ionization and Plasma Electronics", Wiley-Interscience, New York, pages 209 to 217 (1971)).
The frequency of the avalanche pulses rises spontaneously with increasing voltage (see FIG. 1B). This is different from the oscillating electric field employed by the prior art in which the frequency is fixed artificially and remained the same unless human intervention changed it. This difference is at the heart of the present invention because it is not oscillation of the electric field which produces the desired results but these spontaneous, organized avalanches of ion-electrons produced between the electrodes 11 and 12 in air which elicit the Molecular Impulse Response.
Use of a pure DC power supply, with no AC ripple, results in electron avalanches with significantly less pulsing and regularity. Exposure of seeds to these pulses results in a lower seed performance than those exposed to a DC power supply with an AC ripple. Furthermore, results are difficult to consistently reproduce when an AC ripple is absent. Thus it is not merely exposure to an electric field which produces the beneficial results claimed here, nor is it exposure to any type of electron avalanches. The seeds must be exposed to the sharp, regular, uniform or organized electron avalanches as shown in FIG. 1B for best results.
B.) Diagnostic Procedure: Different voltages (generally between 2-20 kV) and different time exposures (from seconds to minutes) produce the best results with different varieties of seed. The optimal parameters are selected for each seed by exposing them at a range of voltages for a range of times, and comparing the results by germination and/or growth and/or yield tests, as well as by redox measurements.
A redox diagnostic procedure allows the achievement of significant improvements in a wide variety of seed/plant types. This diagnostic procedure is necessary because a variety of seed which is positively effected at a high (20 kV) or low (5 kV) voltage may be effected negatively by a medium (15 kV) voltage. Conversely, seeds which do well at a low voltage may do poorly at a high voltage and vice versa.
It has been found that the seeds should be stored at 40 DEG F. to 80 DEG F. If the temperature is too low then no result is achieved.
It will be appreciated that the seeds can be positioned on a non-conductive screen 22, such as fiberglass, between the electrodes 11 and 12 as shown in FIG. 5. Preferably the electrodes 11 and 12 are round with rounded edges. The electrode preferably has a 8 to 9 cm gap and a diameter of about 30.5 cm. The seeds are placed on the electrode so as not to be touching significantly.
EXAMPLE 1
This Example shows laboratory germination tests accurately diagnosing treatment levels which produce yield increases, plus examples of how a voltage which is good for one crop produces marginal or decreased yield in another, as compared to untreated controls as shown in Table
TABLE 1
Best Germ. Good Yield@1
Marginal or
Crop Type kV kV Poor Yield
Tomato 5 kV 4, 12, 16 kV
8, 20 kV
Carrot 5 kV 4 kV 12, 20 kV
Soybeans 8 kV 8,12 kV 4 kV
Navy Beans
10 kV 10, 12 kV 6 kV
Bi-Color 15 kV 16, 8 kV 12, 4 kV
Sweet Corn
Kandy-Krisp
15 kV 16, 12 kV 4, 8 kV
Sweet Corn
Inbred 4, 16 kV 8, 12, 20 kV
Field Corn
Hybrid 4, 12, 16 kV
8, 20 kV
Field Corn
Cypress 15 kV 16 kV
Rice
@1 Measured by fruit and grain weights.
Frequently, laboratory germination voltages were tried in increments of 5, i.e. 5, 10,. 15 kilovolts, while field tests were in increments of four kilovolts, thus producing non-exact matches. Results of a range of treatment durations have been averaged here for each voltage for simplicity.
A key element of the present invention is a waiting period during which treated seeds are not germinated for a minimum of several weeks after exposure. Germination of exposed seeds before this waiting period is completed can result in no improvement in the seeds or even negative effects. Consistent, reproducible, improvements are not found with seeds planted soon after exposure. Improved effects in treated seeds have been seen as long as 18 months after treatment. There is not as yet any known upper limit to the waiting period. While the minimum waiting period varies from one seed variety to another, a minimum of 30 days has been found to be effective. The seeds of FIGS. 2A to 2C were stored for 35, 35 and 36 days respectively.
The redox ratio is a measure of temporal variations in respiration as measured by changes in oxidation/reduction activity in seedlings grown from treated seeds. Increased phase amplitudes of redox cycles, indicative of increased rates of respiration and free radical activity, have been consistently measured in 10-12 day seedings grown from MIR-treated seeds (FIGS. 3A and 3B). Many studies have suggested that alterations in redox ratios are linked with growth responses in biological organisms. (Levengood, "Bioelectrochemistry And Bloenergetics, 19 461-476 (1988); also Allen and Balin, "Free Radical Biology and Medicine" Vol. 6, pp. 631-661 (1989); A. Sakamoto et al., FEBS Letters, Vol. 358 pp. 62 (1995)). Whether or not this is in fact the mechanism of the present invention, alterations in redox ratios have been seen to be linked with improved growth performance in MIR-treated seeds, including eventual increases in final yield. In the green seedling autotrophic stage, redox levels of seedlings grown from MIR-treated seeds are lower than in untreated seedings as shown in FIG. 4, consistent with the hypothesis of higher levels of anti-oxidants present which deactivate free radicals and thereby lower redox ratio levels.
Measurements were made according to the procedure set forth in Levengood, Bioelectrochemistry And Bioenergetics, 19 461-476 (1988). Detection of the above-mentioned free radical alterations can be used as a means of quality control for MIR operations. This monitoring or quality control can serve as a rapid check that the desired effect is being achieved in the treated seeds, without resorting to time-consuming growing of the seeds. This redox ratio analysis makes commercial scale operations reliable and dependable.
From several hours to several days after treatment, MIR seedlings display raised redox ratios, indicating a burst of free radicals within the cells formed by the impact of the ion-electron avalanches. Seeds experience activation of cellular anti-oxidant defenses and consequently have lowered redox ratios. In dried seeds this process moves slowly, as do all metabolic processes in quiescent seeds. Seeds which have been treated at an effective voltage and for an effective time will, during storage, experience a redox level shift as cellular anti-oxidant defenses, such as Superoxide Dismutase (SOD) and others, deactivate the free radicals. In maize, for example, cells have been known to produce more SOD than needed to disable the free radicals present. Gail L. Matters and John G. Scandalios, "Effect of the free radical-generating herbicide paraquat on the expression of the superoxide dismutase (Sod) genes in maize", Biochemica et Biophysica Acta 882 p. 33 (1986) observed 54% increases in SOD levels but only a 40% increase in SOD activity, in response to a burst of superoxide radicals. Thus the resulting surplus of anti-oxidants lowers the normal levels of free radicals in seeds and in mature, developing plant tissue the MIR treated plants have lower redox ratio than in the untreated controls as shown in FIG. 4.
As shown in FIG. 5, the spatial drift of the MIR pulses outside the electrodes 11 and 12 can be examined by stationing an experimental probe coil 101 near the electrodes 11 and 12. A linear chart recorder 21 is used to detect the induced current in coil 101. The electron avalanches drift laterally from between the electrodes 11 and 12 and through an electrostatic-magnetic coupling induce a magnetic field in the coil 101, which in turn generates a potential in the millivolt range. With the coil 101 placed directly across one channel of a dual channel chart recorder such as recorder 21 in FIG. 1A and the MIR system across the second channel, one can examine the effectiveness and form of the pulses in action. For example, the set of curves in FIG. 6 show the magnetically induced and MIR pulses from the coupled system. The coil 101 usually has 10,000 to 100,000 turns, preferably 80,000 turns.
As pointed out by H. Raether ("Electron Avalanches and Breakdown in Gasses", Butterworth & Co. Ltd., U.K. 1964) one reliable criteria to know whether an observed current pulse can be identified with an avalanche process is to compare the form of the avalanche pulse with the induced magnetic component. From the basic theory of electron avalanche formation one should find that the induced magnetic component H (expressed here as coil 101 potential) is directly related to ln(i), where i is the amplitude of the avalanche current pulse in the MIR system. The experimental data in FIG. 7 confirms (r=0.89; P<0.05) that these are electron avalanches.
EXAMPLE 2
When the above steps are used together as part of a coherent process to treat the seeds in the aforementioned manner, the following results have been achieved in a variety of crops in both laboratory and field tests:
1) Increased rate of field emergence. An example is shown in FIG. 8A for Glycine max. Var. PS-202 and in FIGS. 8B and 8C for two varieties of Zea mays sweet corn.
2) Increased rates of plant growth and plant size uniformity.
EXAMPLES 3 AND 4
Examples of the MIR effect in sweet corn are disclosed in Table 2 and 3 below. The data were taken at 52 days development within field test plots. The seeds were stored for 56 days.
Variety-"Kandy
TABLE 2
Plant heights (cm)
N- Coeff.
kV-
Series ave. sd plants of Var.
level
Controls 113.2 29.8 49 26.3% None
5 sec. 145.2 11.3 31 7.8% 12-16
10 sec. 134.8 26.7 37 19.8% 12-16
Variety
TABLE 3
Plant heights (cm)
N- Coeff.
kV-
Series ave. sd plants of Var.
level
Controls 109.6 36.3 81 33.1% None
5 & 10 126.6 28.4 43 22.4% 12-16 sec.
5 min. 123.2 28.4 36 23.1% 12-16
EXAMPLE 5
Increased lateral root growth which has been achieved.
Navy bean seed were treated on Sep. 30, 1992 and germinated 65 days later (20 seeds per lot) as shown in Table
TABLE 4
3 Day
Voltage Duration Roots sd Number
5kV 25 sec. 6.26 cm 1.64 20
10 kV 25 sec. 6.63 cm 0.92 19
Control 0 4.54 cm 2.63 20
EXAMPLE 6
Accelerated maturity has been achieved. Some plants grown under open field conditions from treated seed reach the harvest stage in significantly fewer days, as compared to controls. With sweet corn of two varieties, ears with protruding silk were counted 52 days after they were planted as shown in FIGS. 9A and 9B.
EXAMPLES 7, 8, 9, 10, 11, 12
Increased Yield has been achieved in a variety of commercial crops under normal field conditions, with no extraordinary use of sprays, irrigation, or fertilizer. These effects have been noted in various plants. Soybeans: with a +28.6% increase in yield by dry weight of Soybean seed (Glycine max) of variety 05-202, were exposed for 5 minutes to voltages of 5, 10, 20 and 30 kV on Mar. 2, 1994. One row of 48 seeds from each of these series was planted May 27, 1994 (25 days later) in individual field test plot. Emergence was noted as shown in FIG. 8A, with significant improvements over controls. The best emergence was seen in the 5 kV and 10 kV exposures. These two exposures were the same ones which showed increases in yield at harvest. The results are shown in Table
TABLE 5
Series Voltage Yield in Lbs.
Control Controls 1.75 lbs.
A 5 kV 2.25 lbs.
B 10 kV 2.20 lbs.
D 20 kV 1.63 lbs.
E 30 kV 1.50 lbs.
Soybeans: In a 1995 field test, seeds of Soybean var. "Young" were treated Mar. 15, 1995 and planted May 12, 1995. Each field plot entry represents the mean of four replicates from a two pound lot of treated seed. Results were converted to bushels per acre. Weights per 1,000 seeds from harvest showed appreciable differences. Yield increases were the result of more soybeans produced. The results are shown in Table
TABLE 6
TREATMENT BUSHELS/ACRE
Control 35.95
4 kV, 10 sec. 37.04
4 kV, 30 sec. 34.99
4 kV, 5 min. 36.04
8 kV, 10 sec. 40.10
8 kV, 30 sec. 41.44
8 kV, 5 min. 41.73
12 kV, 10 sec.
34.74
12 kV, 30 sec.
39.50
12 kV, 5 min. 39.64
Control 34.92
Field Corn: 24 seeds per lot were planted on May 31, 1995 in Blissfield, Mich. Figures are pounds of shelled corn per lot. The results are shown in Table
TABLE 7
Inbred, Variety 305-10Gr (F6)
VOLTAGE 10 sec. 30 sec. 5 min.
Control
4 kV 2.65 lbs.
1.85 1.55 2.10
8 kV 1.80 1.95 1.45 1.95
12 kV 1.95 1.35 1.50 1.90
16 kV 1.60 1.00 0.95 2.00
Mean of Controls: 2.03
Hybrid, Variety HYPOP.2830MF. The results are shown in Table
TABLE 8
VOLTAGE 10 sec. 30 sec. 5 min.
Control
4 kV 7.15 lbs
7.10 6.65 5.55
8 kV 5.05 4.40 4.75 4.90
12 kV 5.95 5.65 4.85 4.20
16 kV 5.20 5.95 5.10 6.10
20 kV 5.20 4.75 3.95 3.20
Mean of Controls: 4.79
Carrots: Carrot seeds of variety Daucus carota Danvers 126 were planted May 31 1995 at Blissfield, Mich. and harvested Sep. 7, 1995. Weight per carrot figures are summarized by voltage in FIG. 10. Below are results per treatment duration for 4 kV and 8 kV (best yielding voltages) plus controls. In these results the interplay and dual importance of both time and voltage level is obvious. Here the increases over controls follow no linear progression, emphasizing the importance of the diagnostic procedures discussed earlier in order to select the most effective voltage and treatment duration for a particular seed variety. The results are shown in Table
TABLE 9
VOLTAGE DURATION WT./CARROT
4 kV 10 sec. / 0.10 lbs.
4 kV 30 sec. / 0.112
4 kV 5 min. / 0.141
4 kV 30 min. / 0.128
8 kV 10 sec. / 0.066 lbs.
8 kV 30 sec / 0.154
8 kV 5 min. / 0.175
8 kV 30 min. / 0.093
0 0 0.10 lbs-- /Control
0 0 0.096-- /Control
0 0 0.105-- / Control
0 0 0.089-- / Control
0.098 Mean of Controls
Tomatoes: Seeds of Lycopersicon esculentum variety malinta were exposed Mar. 10, 1995 and planted May 31, at Blissfield, Mich. and harvested Sep. 5, 1995. Yield in pounds of fruit per plant was averaged for each voltage across four time exposures (10 sec., 30 sec. 5 min., and 30 min). The results are shown in Table
TABLE 10
VOLTAGE LBS./PLANT % CHANGE
Control 0.516 / 0%
4 kV 0.69 / +34%
8 kV 0.455 / -12%
12 kV 0.648 / +26%
16 kV 0.61 / +18%
20 kV 0.458 / -11%
Rice: Cypress rice (Oxyza sativa) seed of variety Lemont was obtained from Mississippi State University, treated Mar. 12, 1995, and planted May 11, 1995 (59 days) in Mississippi. Test plots were flushed with water May 15 due to extreme dryness. Emergence occurred May 25 (delayed due to dryness) and plots were flooded June 9. Each figure is the result of 250 gms. of seed grown in four replicated plots, averaged and extrapolated to bushels per acre. Peak yield increases were noted as shown in Table
TABLE 11
VOLTAGE TIME YIELD % CHANGE
Control 0 159.37 / 0%
16 kV 10 sec. 180.13 / +13%
16 kV 30 sec. 169.06 / +6%
8 kV 5 min. 170.08 / +7%
FIGS. 11, 12 and 13 show the circuit 200 of the apparatus of the present invention. The apparatus is available from Hipotronics, Inc., Brewster, N.Y. There is an AC circuit 220 and a DC circuit 240. The negative terminal 260 is connected to the cathode electrode 12 and the positive terminal 280 is connected to the anode electrode 11. The various elements in the apparatus of FIG. 11 are shown in Table
TABLE 12
220 Circuit
C1 .022 600 V
C2 .022 600 V
PLI
F2 2A
UP1
MDV1 250 V
200 Circuit
NE1
NE2
POS Positive
NEG Negative
R1 5 K 1/4 W
R2 5 K 1%
R3 250 K 1%
R4 270 K
A2 Meter Circuit P/N 30-293
C1 .22 400 V
C2 .22 400 V
201 Circuit
T1 Transformer
R1 250 M, 6 W
R2 250 M, 6 W
R3 50 K, 50 W
R4 50 K, 50 W
R5 200 M, 6 W
R6 22 M, 1 W
R7 22 M, 1 W
CR1 Diode
CR2 Diode
C1 0.02 .mu.f; 30 kV
C2 0.02 .mu.f; 30 kV
POS Positive
NEG Negative
Output 60 kV DC
2.5 mADC
FIGS. 15, 16 and 17 show the results of aging of the seeds for a period of time. As can be seen the aging is very important.
FIG. 18 shows the results when oat seeds are treated in the panicle which tends to shield the seed from the electrons. As can be seen, the treatment is effective but less so than in FIG. 17.
It is believed that the influence of the MIR process on seeds is based on the formation of electron-ion avalanches in air at normal atmospheric pressure and temperature. Under an applied electric potential, these avalanches can be directed as electron-ion impulses in the form of regular cycles or plasma waves. The frequency, amplitude and confinement of these pulses are governed by the applied potential and the design configurations of the MIR apparatus.
In the MIR process there is a relationship between the electron-ion avalanche pulse formation and the manner in which they form an organized plasma. The avalanche formation takes place between parallel plate electrodes 11 and 12 at a potential sufficient to cause the electrons (e@-) leaving the cathode to gain enough energy to ionize air molecules through both elastic, and to a lesser degree, inelastic collisions. In the present MIR configuration the minimum potential for avalanche formation is around 0.5 KV/cm. In the electron-molecule collisions new e@- 's are formed and these plus the primary e@- keep repeating this process thus forming a cascading avalanche.
The mean number (n) of drifting electrons e@- 's grow at,
n(x)=exp (.alpha.x) (1)
wherein x is the distance of e@- drift, and .alpha. the mean number of ionizing collisions per e@- per cm. Nasser (E. Nasser, Fundamentals of Gaseous Ionization and Plasma Electronics, Wiley-Interscience, New York (1971)) points out that after a time t' the electric field disappears within the avalanche so that the e@- swarm stops and attaches to molecules, that is, the plasma pulse is partially neutralized or discharged. This takes place inside the electrode gap if the drift path L of the avalanche is,
L=vt' (2)
wherein v, the e@- drift velocity is less than the electrode spacing distance d (in air, v is around 10@7 cm/sec.). With d=8 cm, t' must be <8.times.10@-7 sec. The positive ions (not shown in FIG. 1A) have a low v@+ of around 10@5 cm/sec and therefore have drifted very little from their point of production.
The current i produced by an avalanche is,
i=(.epsilon.n0 /t')exp (.alpha.v't) (3)
If we take (.epsilon. n0 /t') as the rate constant k', for the avalanche formation,
i=k'exp (.alpha.v'T) (4)
where T is the transient time for one avalanche pulse, therefore
ln(i)=k(.alpha.v'T) (5)
wherein k is a new rate constant. Thus in (i) is proportional to the mean number of ionizing collisions (.alpha.) during an avalanche pulse of transient time T.
One reliable criteria (H. Raether, Electron Avalanches and Breakdown in Gasses Butterworth & Co., Ltd., Great Britain (1964)) to know whether an observed current pulse can be identified with an avalanche process is to measure and compare the growth of e@- 's with the theoretical relationship.
n=exp (.alpha.v t) (6)
In the MIR system there is no e@- confinement, therefore the avalanche pulses drift laterally outside the confines of the parallel plate electrodes. This external drift of plasma provides a method for experimentally examining the growth of electrons as predicted by the Equation-6 theoretical relationship. For this purpose an experimental probe coil 101 consisting of 80,000 turns of #40 copper wire, was positioned in proximity with the MIR system (FIG. 5). When placed directly across one channel of a linear chart recorder, any induced magnetic field is readily detected as a voltage pulse in the probe coil 101. Avalanche pulses of varying current amplitudes were formed within the MIR system and recorded on a separate recorder channel as shown in FIG. 6. Any induced field in the probe coil is taken as being proportional to the plasma density formed by the ionizing collisions. From Equation 5 the predicted relationship between a transient avalanche current s and the magnetic field H, induced by an ion-electron concentration (.alpha.) drifting across the test coil 101 would, under these hypothetical conditions be given by,
H=c1 ln(i)+c2 (7)
wherein c1 and c2 are proportionality constants.
From chart recorder traces taken from experiments conducted over a range of electrode potentials, the amplitudes (in mv) of the plasma induced magnetic fields were compared with the amplitudes of the avalanche currents. These data (FIG. 7) plotted according to Equation 7 show good correlation (r=0.89; P<0.05) between the theoretical model of plasma avalanches and the experimental data obtained from the MIR system.
At a given potential the amplitudes and frequency of the avalanche pulses remain relatively constant over the transient intervals. The stability of the ion current pulses was examined by "injecting" excess electrons into an MIR system during a succession of stable avalanche pulses. If UV radiation is directed onto the cathode plate, electrons are released through the photoelectric effect. This can produce what has been called (H. Raether, Electron Avalanches and Breakdown in Gasses, Butterworth & Co., Ltd., Great Britain (1964)) "Avalanches With Successors". Through the injection of additional secondary electrons the amplitudes of the avalanche pulse currents are increased.
This photoelectric avalanche enhancement was produced in a MIR. system consisting of "Optical Transmitting Electrodes" or OTE's (glass coated with a semiconducting tin oxide film) as electrode 12 arranged with electrode separation of 6 cm and 20 kV applied potential. As shown in FIG. 14, the effect of the electron injection is shown to take place 30 seconds after the start (indicated by arrow) of cathode exposure. Due to a shielding effect (E. Nasser, Fundamentals of Gaseous Ionization and Plasma Electronics, Wiley-Interscience, New York (1971)), a plasma will tend to remain stable even when external charges are introduced into the avalanche system. This initial delay followed by a rise to a maximum current amplitude at around 70 sec. followed by the gradual decline, is very consistent with the results obtained in other plasma systems, again confirming that it is a plasma electron avalanche process at work in the space between the electrodes. Exposure of the anode (polarity reversed) to UV had no effect (lower curve) on the current pulse amplitudes, as would be expected. using an anode which is wider than the cathode alters the shape of the electric field in a manner which contains more of the ion/electrons between the electrodes, allowing fewer to drift outside. The result is even more uniform and regular pulses of ion/electron avalanches.
The commercial advantages of the present invention are:
(1) Germination and Early Growth: With the MIR method the plant moves through the vulnerable, seedling stage faster. Greater uniformity at this stage limits the disadvantages of taller plants shading shorter ones and increases chances for all to thrive. Uniformity of growth also makes it easier to harvest the plants.
(2) Root Growth: The MIR method is of particular value in plants such as navy beans where root growth is frequently a problem.
(3) Accelerated Maturity: Accelerated maturity due to the MIR method is of economic advantage to farmers in crops, such as tomato and sweet corn, where the first produce to market each season commands much higher prices. In countries which double crop, it increases the likelihood that both crops will be able to mature and produce a full harvest. In far northern regions, with limited daylight and warm days in growing season, the MIR method increases the chances of a successful season.
(4) Increased Yield: There are economic and humanitarian advantages to the MIR method. There is commercial appeal to the farmer, allowing him to grow more crop to produce income from the same farm. With world population growth outstripping food supply, any significant increases in yield is beneficial.
Key features of the MIR method are:
(1) Sharp, well-organized, uniform electron avalanches (not corona discharge, and not static electric fields). This is provided with a DC voltage source having an AC ripple.
(2) Voltage potentials are 0.2 vK/cm to (but not including) dielectric spark gap breakdown discharge.
(3) Anode electrode with the seeds.
(4) Special electron feedback loop 15 enhances results.
(5) Diagnostic Procedures.
(6) A waiting period of several weeks between treatment and planting.
(7) Redox ratio measurement provides quality control after treatment by the MIR method to confirm if effect was achieved, thus providing an immediate check on results.
(8) Coil 101 recorder system provides an additional quality control to insure avalanches are in fact being produced, and have the proper form. Without this test, humidity and dust/debris on electrodes 11 and/or 12 could cause failure to produce avalanches (particularly when operating near the 0.5 kV/cm threshold, which is frequently used with some seeds.
(9) The MIR method is practical and affordable for large scale commercial operations. Short time period of treatments are required (seconds to minutes) and small amounts of electricity are expended. The MIR method is suitable for conveyor-driven seed handling systems. The MIR method produces consistency of results.
It is intended that the foregoing description be only illustrative of the present invention and that the present invention be limited only by the hereinafter appended claims.
Seed Science and Technology (Netherlands) v. 9(2) p. 567-576 (1981)
Steere, W.C., Levengood, W.C., Bondie, J.M.,
Agro Sciences Inc., Ann Arbor, Mich. (USA),
An electronic analyser for evaluating
seed germination and vigour.
The electrical currents conducted through exudate solutions from 100 individual seeds were measured with a Model ASA-610 Seed Analyser and compared to standard laboratory germination and certain aspects of seed vigour. Examination of the histograms of current distribution from 100 seeds showed a shift towards higher current ranges as seed lot quality declined. A simple technique of data analysis of the current levels provided a "germination prediction" which correlated well with standard laboratory germination. Results from various commercial lots of soybeans (Glycine max), bushbeans (Phaseolus vulgaris), and cottonseed (Gossypium hirsutum) showed that the method has broad potential usefulness as a rapid indicator of commercial seed lot quality.
A method for increasing the proportion of mutants in a generation in a first plant species having a recognized and established phenotype involves the simultaneous somatic exposure of germinal plants of the species to contact with whole cells and associated material of a second species of plants, and to electrophoretic conditions. The plants of the first species are preferably in a germinal state, such as seeds or seedlings, while the whole cells and associated materials of the second species can be a seedling root tip, a seedling, a tissue macerate (suspended in either water or agar) root nodules, fruit tissue or root tissue. When the cells of the first and second species have different membrane potentials, the step of electrophoretic exposure can be carried out by simply placing the cells in contact with one another. Preferably, however, an electropotential difference such as a constant DC voltage is disposed across the somatic cells of the first species of the plant and the whole cells and associated materials of the second species of plant, for example, by attaching one of a cathode and anode to the first species of plant, and the other of anode or cathode to the second species of plant.
BACKGROUND OF THE INVENTION
1. Field of the Invention
The present invention relates to a method for inducing mutations in plants, and more particularly to a method for increasing the number of plants of a first species which exhibit a phenotype or characteristic normally associated with a different species.
2. Description of the Prior Art
The members of a given species of plant typically share a number of well-established physical characteristics associated with the genetic materials of their cells; these characteristics are known as phenotypes. However, it is well known that plants of a given species having one or more new and distinctive characteristics, generally referred to as sports or mutations, occur naturally as a small fraction of any plant population. For centuries, mutants have been selectively bred to produce new varieties or modifications of existing plants. In natural populations of plants, however, the frequency of mutations is generally considered to be less than 1 in 500,000, so that the selection of desirable mutants after such breeding is a slow and laborious process, particularly since it is well recognized that mutants exhibiting a desirable phenotype are rare, and progeny outputs are often low.
Several methods for increasing the occurrence of mutants in a population of a given species are well known; for example, the exposure of such a population to ionizing radiation. Such techniques, however, are typically subject to the drawbacks that the individually resulting mutants are generally weak, and must still be subjected to the time-consuming and labor-intensive techniques of isolation and selective breeding for a large number of generations, before a sufficient number of mutants possessing the new phenotype are obtained for use in outcrossing or agricultural growth.
Recombinant DNA and protoplast fusion techniques are potentially useful for producing new varieties of plants without isolation of mutants or selective breeding. The use of these techniques is subject to several drawbacks, however. First, these techniques are tedious and slow, requiring elaborate instrumentation involving a large number of chemical processes, and a substantial investment in the education and training of the personnel conducting the procedures. Presently, these techniques are very expensive and time consuming. Indeed, Applicant is aware of no reported instance of the inducement of a functional expression of a novel gene (phenotype) from one species of plant to a population of another species of plant, employing these genetic engineering techniques.
SUMMARY OF THE PRESENT INVENTION
The present invention overcomes these and other difficulties encountered in prior methods of inducing mutations in a population of a first species of plant by providing a method for increasing the number of mutants exhibiting altered phenotypic characteristics, characteristics which are stable in successive generations, where such phenotypic characteristics are an established trait of a second different species of plant. The method of the present invention allows for the production of large numbers of plants having substantial modifications from the parent generation, without the delay of several generations for selective breeding and establishment of characteristics as stable by outcrossing, and which does not require the complex instrumentation or large numbers of chemical reactants and steps inherent in present recombinant DNA or protoplast fusion techniques.
The method according to the present invention involves placing a plurality of germinal plants of a first or recipient species, this first species exhibiting at least one established phenotype, in contact with the whole cells and associated materials of a second species of plant, while exposing the germinal plants of the first species to electrophoretic conditions, such as an ionophoretic current. The germinal plants are grown to adult plants, or to a stage sufficient to observe any changes from the established phenotype. The exposure of the germinal plants of the first species to electrophoretic conditions can be carried out by simply abutting a portion of seedlings of the first species with seedlings of a second plant species, when the cells of the first and second species have differing membrane potentials. This can be carried out by excising complimentary sections from the root of seedlings of the first and second plant species, and abutting the cut surfaces of the roots. Preferably, however, an external DC current is applied across the germinal first species plants and whole cells and associated materials of the second plant species by attaching an anode to the plants or materials of one species, and a cathode to the plants or cells of the other species. Typically the plants and materials are exposed to a constant DC voltage having a current density in the range of 10 to 100 microamps per centimeters applied at a potential difference of from 1 to 50 volts for periods of five minutes to 24 hours. In effect, the donor material of the second species acts as an electrode substrate or base contactable with the seedlings of the first species. The donor material is prepared as either a tissue macerate or as whole tissue. The donor material can be placed on sterile cotton or a filter paper which in turn rest on a stainless steel plate electrode. Most preferably, the acceptor tissue or plants of the first species are exposed at the seed or early seedling stage, typically 24 to 96 hours after germination by placing the root apex in contact with the donor-coated electrode, and the shoot apex, cotyledons or coleoptile in contact with the electrode of opposite polarity.
The method of the present invention is preferably carried out with genetically pure, stable and homozygous inbred varieties of lines as the host or acceptor first species. Such well-established lines were used in all of the examples described below, and are commercial varieties which have been released from university or USDA breeding programs for public use.
After exposure, the test seedlings or germinal plants of the first species, along with untreated controls, are developed to maturity under field conditions or in a greenhouse, depending upon expediency. Typically, alterations are observed in the growth rates and yields of the germinal plants actually treated, depending upon the type of donor and the exposure parameters; however, a stable expression of an altered phenotype is typically not seen until at least the second generation bred from the treated plants. The frequency of inherited, varietal alterations resulting from the present method ranges from 5% to 95% of the test population, typically, depending upon the specific procedure and plant species involved. This is a substantial improvement over the proportion of one in a few thousands or several thousands of cells or plants treated by recombinant DNA and protoplast fusion methods.
Not only does the present method yield a significantly increased proportion of mutants in the treated plants, but a significant proportion of the resulting mutants exhibit an altered phenotypic characteristic which was, in fact, an established phenotypic characteristic of the second or donor species of plant. It is believed that this transferred phenotype results from the transduction of genetically associated cell tissue components and macromolecular complexes from the second or donor species into the intact, somatic cells of the first or acceptor species, in such a manner as to alter the genotype and/or phenotype of the plants of the first species. For this reason, plants treated in accordance with the method of the present invention, or grown from plants treated in accordance with the present invention, are designated by generation with the letter "T". For example, the first treated generation of the first species of plant is described as the T-1 generation, while a second inbred generation grown from the adult plants of the T-1 generation are referred to as the T-2 generation. This designation of generations is intended to avoid confusion with the system of F-1, F-2 and so on, normally employed in conventional plant breeding, when crossing for hybrid vigor.
It is thus an object of this invention to provide a method, by means of electrophoresis techniques, for the production of new plant mutations consisting of types and varieties having altered genotypic and/or phenotypic characteristics, that is simple when compared with the recombinant DNA and protoplast fusion methods known in the art. The methods of the present invention do not require complex instrumentation, nor drastic alterations in cell wall-membrane contiguity, particularly the removal of the cell wall as required by prior techniques, or detailed elucidation of chromosome maps.
Another object of the present invention is to provide a method for the production of new varieties of plants that can quickly yield large numbers of healthy plants having substantial modifications from the parent plants, thus eliminating the delay of several generations and large test populations required in prior selective breeding programs, which have been conventionally necessary before the plants can be used in out-crossing. Both conventional breeding programs and the recombinant DNA and protoplast fusion methods generally produce a low yield of mutants which must be selectively grown and bred for a large number of generations, before a sufficient number of stable plants are available for use in programs for developing plant varieties; in contrast, the production of such stable plant varieties is remarkably more rapid in the present invention.
BRIEF DESCRIPTION OF THE DRAWING
A better understanding of the present invention will now be had upon reference to the following detailed description, when read in conjunction with the accompanying drawing, wherein like reference characters refer to like parts throughout the several views, and in which:
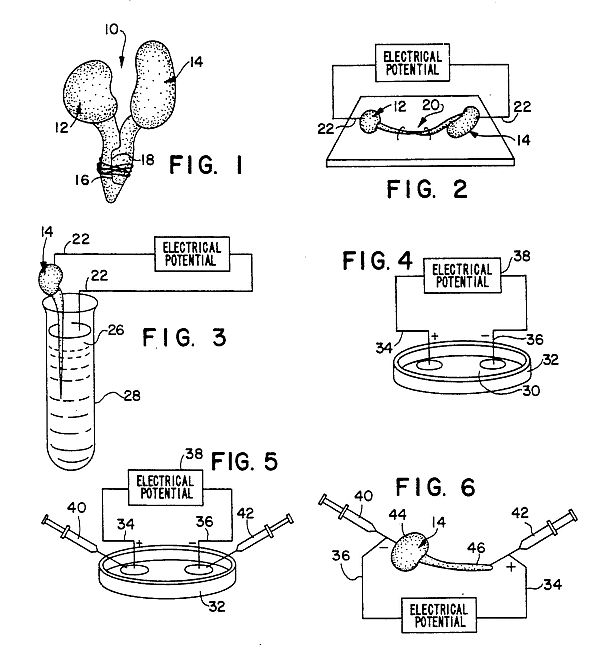
FIG. 1 depicts a joined pair of seedlings of two species of plant;
FIG. 2 depicts a joined pair of seedling with root tips excised and electrodes inserted;
FIG. 3 depicts an electrical potential applied to a seedling in contact with a treated medium;
FIG. 4 depicts the application of an electrical potential to a liquor derived by macerating plant tissue;
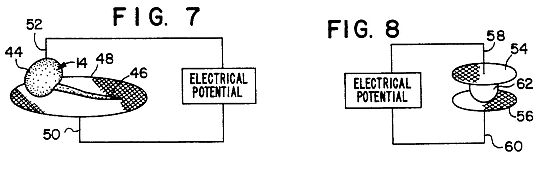
FIG. 5 depicts fractionation of electrode solutions surrounding electrodes;
FIG. 6 depicts the application of an electrical potential across a seedlings and solution filled syringe;
FIG. 7 depicts the application of an electrical potential across a seedling disposed on a treated filter paper;
FIG. 8 depicts a seed disposed between two pieces of treated filter paper with an applied electrical potential;
FIG. 9 depicts the application of an electrical potential across a single cell and a donor medium;
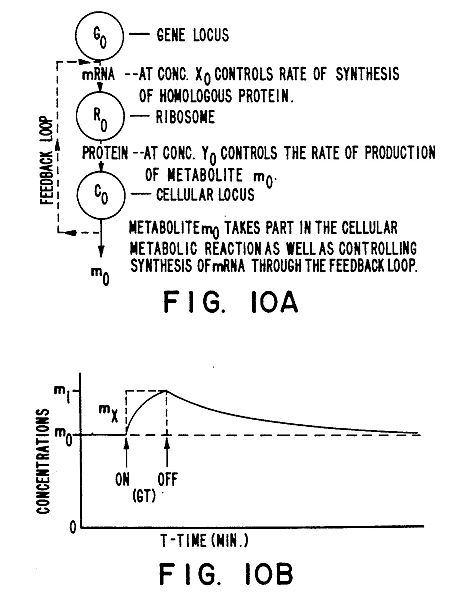
FIG. 10A is a graphic representation of a homeostatic pathway model of the changes induced in plants of a first species by the method of the present invention; and
FIG. 10B is a graphic representation of the response of the concentration of a hypothetical metabolite when a homologous metabolite from a different plant species is introduced into the cell when the method of the present invention is carried out.
DETAILED DESCRIPTION OF THE PREFERRED EMBODIMENTS OF THE PRESENT INVENTION
The method of the present invention both increases the proportion of mutants in a generation of a first species of plant (the species having at least one established phenotype) while simultaneously causing at least some of the resultant mutant plants to exhibit a phenotype, characteristic or trait of a second species of plant. Several illustrative techniques and specific examples of the present invention are described hereinafter. It should be understood that the technique of the present invention is generally intended to be used on a substantial number of plants sought to be modified, so as to provide a ready supply of mutant plants for subsequent varietal development. The electrophoretic techniques described however, can also clearly be conducted on a single plant cell as the acceptor, employing micromanipulative techniques in order to apply an ionophoretic current across the single acceptor cell and donor material. Such a technique is, of course, within the scope of the present invention. In such a case, the acceptor electrode can comprise a thin needle or wire inserted into or in contact with the acceptor cell. In all cases, however, the cell wall and plasmalemma are preferably not breached by the present method.
It is believed that germinal plants, such as seeds or seedlings about one to five days after germinations, are most susceptible to successful treatment by the method of the present invention. While some variation may occur in the percentage of mutant plants grown from the treated seeds or seedlings, such as percentage varying with the species of plants used and the particular technique employed, the method of the present invention will generally result in a substantially greater percentage of mutations than results when radiation or the like are employed to create mutant plants. Moreover, whereas radiation and the like cause random mutations having widely varying characteristics, in general a substantial percentage of mutants formed by this invention tend to exhibit substantially similar characteristics.
With reference first to FIG. 1, a first embodiment of the method of the present invention is thereshown involving a joined pair of seedlings 10 of two different species of plants. One or more seedlings 12 of first species of plant are prepared by longitudinally excising a section consisting of about one-half of the root's side, to expose a fresh cut surface 16. Preferably, the germinated seedlings include radicles in the range of 1 to 6 centimeters in length, and the radicle tip is left intact when the surface is cut, exposing the procambium, protophleom and protoxylem cells. The root side and tip of a corresponding number of seedlings 14 of a second, different species of plant are excised, the radicle tip as well as the longitudinal portion of the side being removed, to form a cut surface 18. The seedlings 14 of the second species are preferably of similar radicle development as the seedlings 12 of the first species, and the procambium, protophloem and protoxylem of the seedlings 14 of the second species similarly form the cut surface 18.
The cut surfaces 16 and 18 of the two species of plants are then immediately abutted and a thin cord is wrapped or tied about the abutted roots in order to insure good contact between them and maintain them in abutment. The excisions on each of the seedlings 12 and 14 should be complementary in order to maximize contact between the cut surfaces 16 and 18. The joined seedlings 10 are planted and nurtured to adult plants, at which time either seeds from the plants are harvested for growth of a T-2 generation from which plants having desired traits are selected; or the T-1 adult plants are directly selected for desired traits. The former is the particularly preferred procedure in this invention.
In the embodiment disclosed in FIG. 1, the seedlings 14 of the acceptor species are exposed to electrophoretic conditions through the existence of a difference between the natural membrane potentials known to exist about both plant and animal cells, Jaffe, Nature, 256: 600-602 (1977). Although natural membrane potentials are known to be of low magnitudes, generally on the order of 1 to 100 millivolts, the adjacent disposition of cells of different species will result in a mutual electrophoretic process. Because each plant species has its own distinctly characteristic metabolic cycle and timing of activity, the biochemical cycles in plants 12 of one species will likely be at a phase different from that of plants 14 of the second species. Consequently, since at one growth stage the mutual potentials may be complementary and at another stage of development they may be opposed, this can provide a potential gradient quite different from that which the cells of the plants 12 of the first species would experience under normal conditions of development.
Because the plant radicle or root tip is responsible for the production of vitamins and other important enzymes used in the development of germinal plants, the plant 14 having the root tip excised will be acceptor plant, while the plant 12 having the root tip retained will be the donor plant.
Applicant has measured the current density in the region where the cut tissues contact, when abutted as disclosed in FIG. 1. For example, when four day old seedlings from different species such as corn and soybean are paired, the current density reaches a maximum value of about 0.7 microamperes per square centimeter at about 40 minutes after initial abutment, with a very gradual decline over the next 10 hours. In contrast, when seedlings of the same species are paired in a similar fashion, such as soybeam-soybean pairings, the current density is only around 0.01 microamperes per square centimeter, again showing a very gradual decline with time. Typically, even at this low electric potential difference between the seedlings of disparate species, new traits appear in the acceptor plants at about a 5% mutuation level and are often in the nature of phenotypical alterations such as plant shape, size and foliage color. The T-1 generation is then selfed to yield a T2 generation, and the altered phenotypes exhibited by the mutated members of the new generations do not segregate out in succeeding generations.
With reference now to FIG. 2, a second preferred embodiment of the invention is thereshown in which the natural membrane potential difference between seedlings of two different species is augmented or reversed, as desired, by the application of an ionophoretic current across the joined seedlings. More particularly, the root tips of seedlings 12 and 14 of two different species are excised, and the cut portions of the seedlings abutted together. A pair of electrodes 22 are then afixed to the seedlings 12 and 14 generally opposite the abutted root portions, for example, in the shoots or cotyledons. An electrical potential such as provided by a constant direct electrical current is then applied through the electrodes 22 across the pair of joined seedlings 20. The voltage applied to the seedlings will generally range between 1 to 45 volts, and preferably on the order of 1.5 to 22.5 volts, for times of about 5 minutes to 24 hours, and preferably about 5 minutes to about 3 hours. This yields a current density across the region where the cut seedlings abut one another in a range of about 10 to 100 microamperes per square centimeter. Preferably, the direction of current applied is chosen to augment the difference in membrane potential of the cells between the different species of plants. Once subjected to such a potential, the seedling pairs 20 are then separated into individual seedlings 12 and 14, which are separately nurtured to adult plants. The selfed T2 generation from the treated T1 generation plants are then selected for desired traits.
Preferably, the electrodes 22 are constructed from iron, since iron electrodes can be inserted into the seedlings without causing detrimental effects to the seedlings. Other electrodes which are not deleterious to plants can also be used, and stainless steel electrodes are particularly preferred for this purpose.
The donor material employed in the present invention need not be a whole plant or seedling. Instead, as shown in FIG. 3, tissues of a donor species of plant can be macerated, such as by blending in water, in order to produce an aqueous donor liquor. The aqueous liquor is collected and added to a support medium such as agar or gelatin, to produce a treated medium 26. The treated medium 26 is disposed in a test tube or vial 28, or other convenient container, and the root tip of the seedlings 14 of the second species of plant are placed in contact with or immersed in the medium 26. Most preferably, the radicle of the acceptor plant seedling 14 is placed in the medium. Once electrode is contacted with the shoot of the seedling, while another electrode is disposed in contact with the support medium 26. An electrical potential is then applied across the seedling 14 and medium 26, of the type, time and intensity described in the preceding embodiment. Following the application of this electrical potential, the seedlings 14 are removed from the treated medium 26 and grown to adult plants, which are then either selected for desired traits or are selfed in order to determine which traits in a T2 generation are inheritable and stable.
The use of agar or gelatin as a medium 26 for suspending the aqueous liquor is desirable, but not essential to use of an aqueous liquor of the donor plant cells. In the embodiment shown in FIG. 3, the medium can be replaced by the aqueous liquor itself. Moreover, the aqueous liquor can itself be subjected to an electrical potential prior to its contact with the acceptor seedling 14, as shown in FIGS. 4 through 6. As above, the tissue of the donor species of plant is macerated in distilled water, and the resulting liquor 30 collected. The aqueous liquor 30 is then deposited in a petri dish 32, and a positive electrode 34 and a negative electrode 36 are placed in the aqueous liquid 30. An electrical potential 38 is then applied to the aqueous liquor 30 across the positive electrode 34 and the negative electrode 36. While the electrodes can be constructed from silver, it is preferred that the electrodes are constructed from platinum in order to reduce oxidation of the electrodes, and minimize the effect of the electrode material upon the aqueous liquor 30. Generally a potential of about 5 to 20 volts is applied for a time of about 10 to 30 minutes. Consitutents of the aqueous liquor will migrate towards or away from one or the other of the electrodes 34 and 36, depending upon the charge possessed by the various tissue constituents. As shown in FIG. 5, the portion of the aqueous liquor 30 which is located about the anode or positive electrode 34 (the anode solution) is removed from the remainder of the aqueous liquor 30 by withdrawal into a hypodermic syringe 40. The portion of the liquor 30 surrounding the negative electrode or cathode 36 (the cathode solution) is removed by drawing into a syringe 42.
The syringes 40 and 42 containing the anode and cathode solutions are then inserted into opposite ends of seedlings 14 of the acceptor species, as shown in FIG. 6, and pressure is applied to the syringes 40 and 42 to inject a portion of the anode and cathode solutions to the tissue of the seedlings. For example, the anode solution contained in the syringe 40 can be inserted into the shoot 44 of the seedling 14, while the cathode solution contained in the syringe 42 can be injected into the root of the seedling 14, preferably into the radicle 46. The positive electrode 34 and negative electrode 36 are then connected to the syringes 42 and 40, respectively (opposite to the electrodes from which the syringes collected a portion of the aqueous liquor), the syringes preferably having metal tips to facilitate electrical contact with the seedling 14. A potential difference of about 1 to 50 volts and preferably of about 1.5 to 22.5 volts is applied to the seedling through the syringes for a time of about 5 minutes to 24 hours, and preferably for about 5 minutes to about 3 hours. Subsequent to the application of the potential difference, the needles are removed from the seedling 14, and the seedling 14 grown to an adult plant. A plurality of seedlings are selected for the desired traits in either the T1 or T2 generation, as described earlier.
In another preferred embodiment of the present invention, only one of the electrode solutions needs to be applied to the acceptor species of plant in order to obtain the high proportion of mutations encountered in the present invention. With particular reference to FIG. 7, either of the anode or cathode solutions collected by the syringes 40 and 42 can be applied to a porous medium, such as a filter paper 48. A seedling 14 of the acceptor species of plant is positioned on the filter paper 48 with both its radical 46 and its shoot 44 in contact with the filter paper 48 containing the donor electrode solution. The donor-containing filter paper 48 is placed in contact with a first electrode 50 while a second electrode 52 of opposite polarity is inserted into the shoot 44 of the seedling 14. As above, the polarity of the electrode 50 in contact with the filter paper 48 is opposite to the sign of the electrode 40 or 42 from which the anode or cathode solution was collected. Because at least some of the constituents of the anode or cathode solution will be of the type to migrate towards the electrode opposite in sign so that of the electrode 50 in contact with the filter paper, these constituents will tend to migrate towards the second electrode 52 upon the application of the potential difference across the electrodes 50 and 52, and thereby across the seedling 14. The length of time and type and strength of potential difference applied across the seedling 14 are as disclosed above. Subsequent to the application of the potential difference, the electrodes 50 and 52 are removed from the seedling, and the seedling 14 grown to either the T-1 or T-2 generation, and selected for any desired traits. The electrodes 50 and 52 are preferably constructed of iron or stainless steel, because of their minimal effects on biological systems.
It should be evident that the embodiments disclosed in FIGS. 3 and 7 are readily adaptable to use in exposing a single cell or isolated protoplast cell of an acceptor species to the aqueous liquor or cathode or anode solutions from the donor species. More particularly, in FIG. 9 there is disclosed another preferred embodiment of the present invention in which a pair of non-reactive electrodes 70 and 72 (preferably platinum electrodes) are used to place an electrical potential across a single plant cell or isolated protoplast 74. The cell 74 is carried on the end of a glass tube 76, the tube 76 being filled with water 78 or another conductive liquid so as to permit manipulation of the cell 74 within the tube 76. The use of a water-filled tube to carry a single plant cell is, of course, a known micromanipulative technique. One of the electrodes, for example, the cathode 72, is electrically connected through the tube 76 and disposed in contact with the liquid 78 in the tube 76. The other of the electrodes, for example, the anode, is electrically connected through the wall of another glass tube 80 and disposed in contact with a donor medium 82 contained in the tube 80. The donor medium 82 is the same as the media prepared in accordance with the preceeding embodiment of the invention. Pressure is applied to the medium 82 to express a small droplet 84 of the medium 82 out of the end of the glass tube 80. The tubes 76 and 80 are mounted to a micromanipulator (not shown), which aligns the tubes 76 nd 80 and permits the droplet 84 to be brought into contact with the plant cell 74. Alignment and contact can be visually monitored through a microscope 86. The plant cell 74 is then subjected to electrophoretic conditions by the application of a DC voltage across the electrodes 70 and 72. The applied voltage should be sufficient to produce a current density in the range of 1.0 to 100 microamps per square centimeter, for a time of about minutes to three hours.
A final preferred embodiment of the general method of present invention is shown in FIG. 8 in which a seed 62 of an acceptor species of plants id disposed between two pieces of porous material or filter paper 54 and 56. Cathode and anode solutions of a tissue macerate of a donor species are prepared as described above. The filter papers 54 and 56 are placed on electrodes 58 and 60, and infused with the anode or cathode solution collected from the electrode 34 or 36 of potential opposite to the electrodes 58 and 60. One of the filter papers 54 or 56 is placed in contact with the hilum or embryo end of the seed 62. The electrodes 58 and 60 can be constructed of various materials, preferably stainless steel or other iron material. It is preferred that the electrodes do not contact the seed 62 directly. An electric potential is the applied to the electrodes 58 and 60, and thus applied across the seed 62. The potential can be applied to the dry seed 62, or the seed can be allowed to be partially or completely imbibed with water or the anode and cathode solutions, before the potential difference is applied. A constant direct current of 20 to 90 volts is applied to the dry seeds, or a potential of 1 to 40 volts is applied to the partially imbibed seeds, for about 5 minutes to 1 hour. After such treatment, the seed may be returned to the quiescent state and stored until it is convenient to plant them. Alternatively, the seeds may be germinated immediately, sprouted and grown to adult plants. Adults in the T2 generation, selfed from the T1 plants, are selected for desired traits.
The methods of the present invention are further illustrated by several following examples. Some of the examples have been followed through the T5 generation in extensive agricultural testing. In general, it has been found in the invention that the induced mutations recognizably segregate in the T2 or subsequent generations, so that selections for further crossings or further development can accordingly be made in the T2 generation. For the most careful screening of the types of mutations, it has been fund advantageous to examine plant row tests in the T2 generations, that is, to use seeds from the individual treated plants of the T1 generation for inbred or selfed plant row replications, in the T2 generation testing. This allows a more efficient screening and categorization of the induced mutations from the T1 generation since traits or characteristics which are not reproduced in a selfed or inbred generation are neither stable nor of particular commercial value.
The high percentage of mutants obtained in the method of the present invention allows a relatively small number of seeds or seedlings to be treated in the T1 generation (which is also referred to as the transduced series), on the order of 15 to 20 seedlings of each transduction polarity being examined for differences in growth and for phenotypic variations, along with a control group of untreated seedlings or seeds of equal number. Thus, about one-third of the plants field tested at the time of the T1 generation are control plants. After the T1 generation, each of the treated lines and controls are grown in three replicated rows of 40 to 50 seeds each within statistically randomized test plots. Unless otherwise indicated, the Latin Square method of randomization was employed. Subsequent to the T2 generation the lines are selected and expanded according to the apparent importance of the new characteristics of the mutant plants.
The acreage necessary to adequately insure that the new characteristics are stabilized in the particular treated lines will vary according to the percentage of mutants obtained in the T1 generation and the number of lines that appear desirable to investigate. For example, in 1984, applicant produced T1 transduced series of acceptor species including corn, tomato, soy beans and navy beans. Less than a one acre test plot was required for 124 transduction series and controls. These particular tests were made in lower Michigan. By 1985, the subsequent T3 generation testing involved an area of 10 acres, while the 1986 T5 generation required over 70 acres of primary growth, in addition to replicated tests at several locations and in several states. The T2 and T4 generations of these transduced series were seed expansion grow-outs in Hawaii, in order to shorten the time necessary to achieve the T5 generation.
The following examples serve to further illustrate the present invention:
EXAMPLE ONE
Longitudinal sections from soy bean (Glycine max) seedling roots (the donor species) were excised in a plurality of seedlings, and longitudinal sections including the root tip were excised from a plurality of bush bean (Phaseolus vulgaris) seedlings (the acceptor species). Each seedling has a radicle in the range of 1 to 6 centimeters in length, and the excised portions were of complementary shape, such as to expose the procambium, protophloem and protoxylem cells of each root tip. The cut portions of pairs of seedlings of the different species were abutted and bound with thread, as shown and described in conjunction with FIG. 1. The pairs of joined seedlings were grown to adult plants.
One in twenty bush bean seedlings so treated resulted in an adult plant that was shorter than the control plants and which has more compact foliage than the control plants, characteristics which are of commercial importance in the harvesting of bush beans. Tall plants tend to lodge and intertwine, and are thus less efficiently harvested. These plants also had leaves of a deeper green color than the control plants, the fruit of these plants and these plants exhibited greater drought resistance than the control plants. Yields under field condition, however, were found to abe about the same as those of the control plants. The seeds of these plants were observed to be intermediate in shape between the soy bean and bush bean progenator seeds.
These new characteristics were stable; they were observed without change through seven inbred or selfed generations with no reversion back to the height, bushiness, color, sweetness, and drought resistance of the original and control bush bean plants. Six generations of the mutated plants, along with an equal number of controls, were grown under field test conditions as described earlier. The maintenance of these characteristics for seven generations demonstrates that the changes were inheritable. The fact that the inbred, transduced plants do not segregate or revert, that is, return to the characteristics of the control plants, demonstrates that the method can provide new varieties of plants which breed true. As will be subsequently discussed, this non-segregating, stable nature of the growth alterations suggests a non-Mendelian or cytoplasmic type of inheritance.
EXAMPLE TWO
Longitudinal sections from a plurality of bush bean seedlings (Phaseolus vulgaris) roots, the donor species, and longitudinal sections including the root tips from soy bean (Glycine max) seedlings, the acceptor species, were excised to expose procambim, protophlem and protoxylum cells on each seedling. Each seedling was germinated and possessed radicles in the range of 1 to 6 centimeters in length. The excised portions of pairs of seedlings of different species were cut in complementary shapes, and the exposed cut portions of the seedlings were joined together to form pairs of joined seedlings in the fashion shown in and described in conjunction with FIG. 1. Each pair of joined seedlings contained a bush bean seedling and a soy bean seedling. The pairs of seedlings were then grown to adult soy bean plants, and one in ten of the soy bean plants so grown exhibited seeds that were intermediate in shape and color between the seeds of the bush bean and the soy bean progenators. The leaves of the one in ten altered soy bean plants were less lobed in shape than the leaves of the control plants, the stem node lengths were reduced as compared to those of the soy bean control plants, and the number of stem nodes was increased as compared to the controls as well. This resulted in a line of altered soy bean plants which had more compact foliage than the control plants and was thus more resistant to lodging under field conditions. These changed characteristics were maintained in inbred or selfed plants grown through four generations. Three of these generations were grown along with an equal number of controls under field conditions.
This example is, of course, the reciprocal or reverse of the transduction which occured in Example 1, that is, the donor and acceptor species are reversed. Significantly, the percentage of altered or mutated plants obtained is of the same order of magnitude in each example, demonstrating that the method allows modifications to be made to plants in two directions. Typically, attempts to induct positive and viable mutations in plants by conventional methods such as by chemical or ionizing radiation treatments yields an expected frequency of useful, viable mutations or phenotypic alterations of one in five hundred thousand test plants (a frequency equal to 0.000002). Examples 1 and 2 demonstrate that the method of the invention can produce plants having new, inheritable characteristics at a rate of 25,000 times that expected under conditions of conventional chemical or radiation treatment. This increase in the rate of mutation is highly significant and commercially valuable in terms of time, space, and the volume of plants needed to be treated or exposed in order to produce positive mutations.
EXAMPLE THREE
Tissue from the immature fruit of tomato (Lycopersicon esculentum) was macerated in distilled water and the resulting aqueous liquor placed in a perti dish. A pair of spaced silver electrodes were inserted in the macerate liquor and a constant direct current electrical potential of 9 volts was applied for 20 minutes. A portion of the liquor surrounding each electrode was drawn into a hypodermic syringe having a conductive needle tip. The conductive syringe tips were inserted into the root and shoot of a plurality of soy bean (Glycine max) seedlings in the fashion shown in and described in conjunction with FIG. 5, 6 and 6, above. A negative electrode was then connected to the syringe containing the solution which has surrounded the positive electrode in the petri dish, while a positive electrode was connected to the syringe containing the other electrode solution. A constant DC electrical potential of 22.5 volts was then applied for five minutes, so that a current of approximately 100 microamps was passed through the seedlings.
Two series of 20 seedlings were treated and grown along with 20 non-treated controls. In one treated series, the electrode solution from the positive electrode in the petri dish was applied to the seedling roots. In the other series, the electrode solution from the negative electrode in the petri dish was applied to the roots of the seedlings. After such treatments, all of the seedlings were grown under field test conditions as described above, and the results obtained are given in Table I below. The asterisk indicates data which is statistically significant at about a 95% confidence level (P less than 0.05). The observed increases in pod and seed yields continued in two subsequent generations of selfed or inbred plants, grown under field test growth conditions.
TABLE I
(N = 20 plants per series)
ELECTRODE SOLUTION
PODS PER PLANT
AVERAGE SEED YIELD
APPLIED TO ROOT
AVERAGE
S.D.
(GRAMS PER PLANT)
ANODE (+) 59.1* 43.6
16.76
CATHODE (-) 49.6* 36.7
14.06
CONTROLS 32.5 13.7
9.50
EXAMPLE FOUR
A portion of root tissue from Eastern Marsh Cabbage Plant (Symplocarpus foetidus) was excised in early Spring (mid-March), macerated in distilled water and admixed with a sufficient quantity of agar to create a donor macerate of moderate viscosity. A portion of this donor macerate was placed in a test tube. The radicles of a plurality of tomato seedling (Lycopersicon esculentum) were immersed in the donor medium. One electrode was inserted into each of the seedlings, while another was positioned in contact with the donor medium. A direct current 9 volt potential difference was applied across the electrodes, and thus across the seedlings and macerate, for five minutes.
Tomato is well known to be one of the agronomic crops which can be commercially grown both under greenhouse and field conditions, while it has been noted that the Marsh Cabbage possesses a high metabolic output in its early stages of growth, R. M. Knutson, Science 186: 746-747 (1974). In view of the hypothetical model set forth in the discussion following the examples herein, and in light of the fact that certain characteristics of the transduced plants in Examples 1 and 2 were intermediate the characteristics of the donor and acceptor species, it was thought there was a significant chance that the high metabolic output of Marsh Cabbage could be imparted to tomato seedlings to increase their fruit yields, and thereby increase the commercial value of the crop.
A number of tomato seedlings so treated were grown in a greenhouse, and the number of plants resulting from treatment, and the number of fruit borne by those plants at the time of fruit ripening, are shown in Table II. Both the positive and negative electrode orientation data were combined in the data reported in Table II, since in this case there were no apparent polarity differences. Again, the asterisk indicates data which is significant at a 95% confidence level (P less than 0.05).
TABLE II
FRUIT/PLANT
DONOR AVERAGE AND s.d. N-PLANTS
Macerate
*4.33 (2.64) 15
Controls
2.43 (2.42) 21
This same donor/host transduction was repeated for the purpose of examining yield levels under field conditions. Using three different varieties, a total of 24 test series were prepared with 30 transduced seedlings in each series (15 per electrode polarity) plus 15 control, non-transduced plants. Exposure was again conducted with the apparatus shown in, and the method described in conjunction with, FIG. 3, at a direct current potential of 9 volts and an exposure time of five minutes. All plants were handles and reared under similar conditions of field environment. Yields from individual plants were recorded at the time of optimum fruit harvest (approximately two-thirds mature fruit per plant).
Of the 24 test series, 7 of them, or 29.2%, disclosed a statistically significant yield advantage (based on mean weight of fruit per plant) over the control or non-transduced groups, at a confidence level of 95% (P less than 0.05). Within the groups showing yield increases, there were also concomitant, statistically significant increases in growth rates and in plant size. The yield data possessing this significance ranged from +35% to +70% fruit weight increases over the controls. An experienced plant geneticist and breeder observing the transduced series selected one with a +50% (P less than 0.05) yield increase as having improved phenotypic characteristics for traits desirable for commercial harvesting, specifically, upright plants having good clustering of fruit.
Individual plants were selected and grown in T2 generation field replications as plant rows (30 plants per row) from this particular exceptional plant series as well as from several of the T1 generation series, including some that showed no yield advantages. Yields were again recorded in the T2 generation. These plants row data disclosed that those plants showing growth and yield advantages in the T1 generation also gave high growth rates and yields in the T2 generation, whereas those showing no growth or yield advantages in the T1 generation gave no growth or yield increases in the T2 generation. From the T1 generation exceptional progenitor plant, a total of 10 plant rows gave statistically significant yield advantages ranging from +40.7% to +55.6%, compared with the T2 generation control plant yields. T3 inbred generations of these high yielding plants are currently being compared in several large scale field tests (approximately five acres at four different locations) with two high yield commercial varieties, as well as with the non-transduced F3 generation controls. In all locations the transduced line is still showing significant growth and development advantages over the non-transduced varieties.
This example illustrates the consistency of the induced phenotypic effects and practical increases in the rates of fruit production in a commercially valuable crop, when the method of the present invention is practiced.
EXAMPLE FIVE
The Symplocarpus foetidus root tissue used as donor material in Example Four was prepared in the early spring (mid-March) growth period, when the metabolic activity in the root was at a high rate. Donor tissue prepared from different tissue regions of the donor plant and taken at a later stage of maturity can have significantly different effects on the growth rate in the acceptor plant Lycopersicon esculentum. Tissues from the root, the development spadix in the lower stem of the plant, and the leaf foliage of the Marsh Cabbage were collected in mid-April, and donor macerates were prepared as described in Example Four and shown in FIG. 3. Seedlings from four different commercial and established varieties of tomato plants were treated with these macerates and with a direct current 9 volt potential and five minute exposure. The tomato seedlings were grown under greenhouse conditions and periodic growth data was obtained. Table III presents data obtained at six weeks of growth which shows the percentage of the total series which possessed growth statistically significantly higher (P less than 0.05) than the corresponding control series.
TABLE III
DONOR TESTS WITH SIGNIFICANT TISSUE GROWTH N-TEST SERIES
Root 2.8% 36
Spadix 19.4% 36
Foliage
30.0% 20
The foliage employed as a donor, with its high rate of protein synthesis, yielded the highest percentage of tests showing significant growth increases in the acceptor series, when compared with the controls. It is noteworthy that the root macerate used in the above test produced significant growth increases in only 2.8% of the series, whereas in Example Four the root tissue obtained about one month earlier (when at its high level of metabolic activity) induced high growth in 29.2% of the test series.
This example serves to illustrate the importance of the selection of tissue for the transduction donor, as well as considering its state of maturity.
EXAMPLE SIX
Many varieties of plants in the pea and bean family (legumes) have the ability to more efficiently utilize or fix nitrogen from the atmosphere than other plants. This diazotrophy occurs through bacteria which live symbiotically on the plant roots and form outgrowths or root nodules. The results of this example suggest that an acceptor species in the cereal family such as corn, which does not fix nitrogen, could have mutations and growth stimulation induced therein from a donor bean species which has these root nodules.
A donor extract was prepared from soy bean (Glycine max) root nodules excised from plants grown from seeds which were initially inoculated with the bacterium Rhizobium japonicum, which is known to produce diazatrophy in soy beans. The macerated nodule liquor was mixed uniformly with agar as a base, and corn (Zeas mays) and sunflower (Helianthus annuus) seedlings were both treated with this donor extract in fashion shown in and described in conjunction with FIG. 3.
EXAMPLE SIX (A)
Both corn and sunflower seedlings were placed in the base medium and exposed to a potential giving an initial current of about 30 microamps through the seedlings. After exposure the seedlings (along with equal numbers of controls) were planted in a field test plot, with no fertilizer added. Growth and development studies were conducted on three separate test series of corn and two separate test series of sunflower seedlings. The growth and development enhancement produced by the root nodule extract treatment was consistently observed in all five test series. Examples of growth and development data are presented in Table IV for a field test series of corn and in Table V for a field test series of sunflower plants. The corn seedlings were exposed to the current for one hour, with the cathode inserted into the donor medium; the data of Table IV were obtained from twelve plants in each series. The sunflower seedlings were exposed to the current for 30 minutes, with the cathode inserted into the donor medium; the data of Table V were obtained from twenty plants in each series. The differences in growth shown in the last column of each table were significant at the 99% confidence level (P less than 0.01).
TABLE IV
DAYS AFTER ROOT NODULE EXTRACT
CONTROLS GROWTH
PLANTING AVE. S.E. AVE. S.D. DIFF.
7 9.25 cm
2.14 cm
5.09 cm
1.64 cm
+81.7%
19 47.00 6.41 31.45
8.89
+49.4%
31 83.58 9.99 64.64
16.83
+29.3%
46 104.92 13.14 80.45
19.44
+30.4%
62 109.00 25.04 74.64
37.37
+46.0%
After 73 days of field growth, the root nodule group disclosed an 84% near development and the control group only a 16% ear development. After 90 days field growth, the average ear weight of the root module series was 55.0 g and the average ear weight of the control series 28.7 g. The kernals on the treated series were also more fully developed than were those of the controls.
TABLE V
DAYS AFTER ROOT NODULE EXTRACT
CONTROLS GROWTH
PLANTING AVE. S.D. AVE. S.D. DIFF.
10 8.00 cm
1.56 cm
6.00 cm
1.81 cm
+33.3%
22 20.30 3.15 16.00
3.89
+26.9%
37 67.00 10.87 56.25
11.18
+19.1%
53 103.10 16.41 86.90
18.17
+18.6%
At maturity the mean seed pod weight (before seed removal) of the nodule-treated group was 21.7% higher than the mean seed pod weight of the control group.
In these field test series, the polarity conditions were limited to the donor medium electrode being the cathode. The reason for examining only the one polarity condition was the fact that preliminary studies with the soybean root-nodule extract disclosed a greater growth response with the donor medium media electrode being negative than with the medium electrode being positive.
This example shows the induction of more efficient growth, development and yield from the root-nodule extracts, as compared to the control plants.
EXAMPLE SIX (B)
Corn seedlings were placed in the donor medium and exposed to a direct current 15 volt potential giving an initial current in the range of 30 microamps through the seedlings. After 10 minute exposures, groups of 15 test seedlings along with equal number of controls were planted in a field test plot, no fertilizer added, and growth data taken periodically during the growth cycle. The test was conducted with five pure and commercially available inbred varieties, Mo17-Ht, A634-Ht, A632-Ht, B73-Ht and W117-Ht, and thus provided range of different lines of stable but homozygous test material. The use of different inbred lines also provided a germ plasm for subsequent hybrid crossing studies.
A total of 18 test series were prepared and examined under field conditions in accord with this protocol. The observed development alterations in the T1 generation were primarily in the rates of maturity or tassel development, in growth rates and in changes in root structure and morphology. The roots in several of the treated series disclosed a much more branching or dendritic patterning, with thickening at the terminus of the root. The roots of the control plants had less branching with no thickening at their termini. This formation of inchoate nodules and alterations in the root morphology of the corn plant is indicative of the initial stages of diazotrophy induction in this cereal plant. Plants from those groups disclosing significant increases in the rate of tasseling or growth were then selected on an individual plant basis for T2 generation self pollination. These T2 general plants were then used in T3, T4 and T5 generations, for both inbred and hybrid crosses.
The advantage of the use of genetically pure, homozygous inbred varieties or lines as the acceptor materials is that mutations in corn can be keyed to alterations in particular chromosomes from known listings. Specifically in the case of corn, new genotypic and phenotypic expressions can be compared with those listed in The Mutants of Maize, N. G. Neuffer, et al Crop Science Society of America (Madison, Wisc.), 1968; and Maize for Biological Research, W. F. Sheridan, Ed. Plant Molecular Biology Association, (University Press, N. Dak.), 1982. It is well known to those skilled in the art that if a particular characteristic appears in a subsequent generation of a plant line where this characteristic was not previously present, a point mutation has occurred on a particular chromosome. Indeed, these point mutations are cataloged in this fashion.
The treatment of corn seedlings by the present invention with the soybean root module donor material produced a number of changes in characteristics in the T2 generation which are known to be associated with particular point mutations. In particular, a number of these mutations are known to be located on chromosome which occurred in the T2 generation plants obtained from some of the treated series. A summary of the frequency of mutations found in two of the inbred treated series (derived from the A632-Ht acceptor) are listed below in Table VI. There were no mutations or phenotypic alterations (a zero percent level) observed in several thousand control or non-transduced plants from this same inbred line.
TABLE VI
CHROMOSOME-3 POINT MUTUATIONS
(T2 GENERATION FROM TRANDUCED INBRED A632-Ht)
MUTATION SERIES M33-1-18
SERIES M33-1-7
Dwarf 7 (20%) 0
Short 7 (20%) 0
Dwarf-Crinkly Leaf
5 (14%) 13 (30%)
rinkly Leaf 0 8 (19%)
Short-Romosa 0 8 (19%)
Dwarf-Crinkly-
0 1 (2%)
Romosa
Normal 16 (46%) 13 (30%)
TOTAL PLANTS 35 43
The probability of any one of these mutations occuring in one plant by change alone is about 1 in 500,000 whereas in Table VI there are shown several cases in which a number of plants expressed two mutations and in one case, a single plant expressed three mutuations. Now, from the laws of strict probability, the odds that these percentages occurred by random chance are, in the case of two mutuations on the same plant, one in 2.5.times.10@11 and, in the case of three mutuations on the same plant, one in 1.25.times.10@17. In addition to point mutuations, other transduced series were observed to express large increases in point mutuations which are known to involve several gene alleles. Examples of these multiple allele mutuations are listed below in Table VII.
TABLE VII
MUTATION TRANSDUCED SERIES VAR-
PLANTS ALBINO LUTEUS IEGATED TOTAL
MED. 27 26% 27% 0 144
(A632-Ht)
MED. 25 0 0 10.7% 28
(A632-Ht)
CONTROLS 0.024% 0.037% 0% 8179
(A632-Ht)
The data of Table VII show a mutuation increase for both albino and luteus of about a thousand times the level observed in the control population. Many of these mutations are not of commercial interest. For example, albino plants do not produce chlorophyll and expire before maturity. However, there were other mutations which have importance in plant breeding. The dwarf plants listed in Table VI are an example of a useful mutation. These plants are about one half the height of the control plants, but the ear size and production were comparable to those of the controls. This normal ear size on the mutuant dwarf plants is an important and commercially beneficial distinction from dwarf corn plants derived from conventional breeding programs, the difference being that the ears on the conventionally bred dwarf plants are small when compared with the normal hybrid ears, and have large areas on the ears which do not develop kernels at all.
Field studies of dwarf plants obtained from corn seedlings treated in accordance with the method of the present invention establish the existence of a number of commercially important characteristics. The following has been shown to be true from five generations of field trials:
The inbred, dwarf mutants have held their recessive characteristics through the T5 generation and exhibit a 50%-60% reduction in plant height, when compared with untreated parent inbred control corn plants, yet produce full ears of normal size, as compared to the controls.
Using this same method and the same soybean root-nodule macerate as the donor material, the dwarf traits have been produced in treated series from four of the five original inbred varieties.
When T5 generation dwarf plants originating from two different treated inbred lines are crossed in a normal manner to produce a hybrid, the dwarf characteristics are transferred to the hybrid. The resulting hybrid is uniformly about 40% of the height of the hybrid resulting from a cross between two untreated, inbred parent lines.
The ear size and kernel formation in the dwarf hybrids are about the same as in the untreated hybrid controls. The commercial significance of this is that a smaller plant size in the dwarf hybrid allows a higher plant density under field planting conditions, which in turn results in a higher yield per acre.
Lastly, the ears on the dwarf plants are located much lower on the plant than on the normal or control hybrids, and thus are more efficiently harvested than those on taller control plants.
Additionally, a male sterile, cytoplasmic mutation (Cms) was observed in 100% of the plants in one of the transduced, Mo17-Ht inbred lines. This mutation is commercially important in the development of inbred lines which do not require the laborious task of de-tasseling in the normal production of hybrids.
In the T3 generation, a number of plants selected for phenotypic growth and yield advantages were used for hybrid crossing studies. In general, the early development and high yield traits present in the T3 generation plants were transmitted into the hybrids when the treated progeny were expressed through the female line of the hybrid. An example of this is a soybean root-nodule donor series expressing the mutuation "prolific", which relates to the percentage of plants with multiple ears. A normal hybrid line has about a 10% level of prolific plants. In hybrid crosses, using female parents from T2 generation inbreds, a direct correlation was observed between the percentage of plants with prolific mutations and the resulting yields. The yields from three field replications were compared with a good producing commercial hybrid. The yield from one of these high producing treated lines is compared with the control hybrid in Table VIII below.
TABLE VIII
PRO- YIELDS
HYBRID SERIES LIFIC (g/plant) YIELD INCREASE
Control (HL2454)
8% 200.09 --
Female Transduced
47% 257.87 +28.9% (P < 0.05)
This example illustrates the number and type of mutations which can be induced by the methods of the present invention. Many of the mutuations have utility in the production of new varieties and in the hybridization of plants. The useful mutant characteristics are selected from the test populations by conventional segregation testing methods commonly employed by plant breeders. The useful mutations are also expressed when employed in hybrid crosses.
EXAMPLE SEVEN
As noted in Example Six, the Eastern Marsh Cabbage (Symplocarpus foetidus) has a high metabolic output during early spring growth, the result of which is development of the plant during a period of temperatures too low for growth to proceed in most plant species. This metabolic response can be imparted to corn (Zea mays) by the method of the present invention, when a donor extract from Marsh Cabbage is applied to the corn seedlings. Potential benefits of such a characteristic might be expressed as higher yields, faster development rates or other useful mutations. A new variety with some or all of these attributes could be grown in regions of the world where the growing season is conventionally believed to be too brief for corn development.
EXAMPLE SEVEN(A)
A donor medium was prepared from the macerated roots of the Easter Marsh Cabbage, and corn seeds were exposed to a direct current during initial inbibition with the medium with the apparatus shown in, and by the method described in conjunction with, FIG. 8. After treatment, the extract-exposed and control series were examined under field growth conditions. Table IX discloses growth data taken just before mid-maturity (36 days after exposure). Each series contained 16 plants. Only the series having a positive base plate polarity during exposure of the seeds exhibited a statistically significant increase (P less than 0.05) in growth, as compared to the controls.
TABLE IX
BASE PLATE PLANT GROWTH PERCENT POLARITY AVE. S.D. CHANGE
(-) 0.878 0.284 +9.5%
(+) 1.003 0.234 +25.1%
Controls 0.802 0.134 --
The polarity differences shown here are consistent with those mentioned in Example Six(A). With the base of the apparatus being the positive electrode, the embryo or radicle end of the seed was disposed upwardly, in contact with the cathode. This arrangement is the one which exhibited a statistically significant increase in plant growth. Cathode-radicle exposure was also the optimum situation for the plant series reported in Table IV and V. This demonstrates the consistency of the electrode orientation in the method of the invention.
A detailed field examination of the plants listed in Table IX disclosed five unique plants out of each group of 16 treated series. Each of these five plants had definite growth enhancement, larger and greener foliage, the foliage being more pronounced than even the other members in the same test series. The growth of these designated "sub-groups" are listed in Table X, again at 36 days after exposure. The differences in growth between the sub-groups and the controls were statistically significant (P less than 0.01).
TABLE X
SUB-GROUP PLANT GROWTH GROWTH
POLARITY AVE. S.D. N-PLANTS DIFF.
(-) 1.208 m 0.039 m 5 +50.4%
(+) 1.238 0.070 5 +54.4%
Controls 0.802 0.134 16 --
Displayed in Table XI are data showing the differences between the leaf blade width in the two sub-groups and the controls. These data were taken at nodes 6 and 7 at 106 days of maturity. The differences in leaf width between the treated and control series are statistically significant (P less than 0.01).
TABLE XI
LEAF
SUB-GROUP MAX. WIDTH WIDTH
POLARITY AVE. S.D. N-LEAVES DIFF
(-) 9.29 cm 0.91 cm 16 +15.6%
(+) 9.44 0.87 16 +17.4%
Controls 8.04 0.79 16 --
Development was also more rapid in these sub-group plants. At 82 days development, both sub-groups disclosed 100% tassel formation, whereas in the controls only 37% possessed tassels. The positive polarity sub-group also disclosed two developing ears, with no ear development at all in the controls.
The final yield results for each entire series of plants is shown in Table XII. The ear weights are somewhat lower than normal, especially in the control series. This was due to a dry period during early ear development, a situation which occurred throughout the Midwest in the 1983 growing season. All three series were, however, subjected to the same water stress conditions. The data in Table XII show the importance of early ear development in the two test series which occurred before the water stress interval.
TABLE XII
BASE PLATE EAR WEIGHT WEIGHT
POLARITY AVE. S.D. DIFFERENCE
(-) 78.5 g 51.3 g +187.6%
(+) 131.4 80.2 +381.3%
Controls 27.3 24.2 --
The final ear weights from the two sub-groups of special high vigor plants gave values of 134 g per ear for five negative base plate polarity plants, and 230 g per ear for the five positive base plate polarity plants. The controls averaged only 27.3 g per ear. The differences are significant at a 99% confidence level (P less than 0.01).
This example demonstrates the induction of a metabolic response having a positive effect on both development and yield in corn, when the corn seeds are treated in accordance with the method of the present invention. It also shows the practical value of selecting outstanding plants in a given test series.
EXAMPLE SEVEN(B)
A donor medium was prepared in mid-March from the macerated roots of the Easter Marsh Cabbage. Corn seedlings were exposed to the donor macerate with the apparatus shown in, and in accordance with the method described in conjunction with, FIG. 3, and with the test conditions described in Example Six(B). The same five inbred lines, also as described in Example Six(B), were utilized. After exposure the treated and control series of plants were examined under field test conditions as outlined in the previous examples.
In the T1 generation, the plant alterations in the treated series of plants were expressed as increased development rates, plant size and plant shape variations. From these treated series of plants, individual plants were selected for T2 to T3 generation inbred and hybrid crosses. In the T2 generation, several point mutations were observed, and their degree of expression is listed in Table XIII, along with the associated allele and chromosome on which the mutuation is known to occur. None of these mutuations was found in several thousand untreated controls.
TABLE XIII
MUTATION
CHROMO- RECENT
NAME ALLELE SOME NO. EXPRESSION
Rust Resistant
Rp 10 100%
Zebra Necrotic
zn 10 10%
Purple pl 6 50-100%
Pigmy pv 6 25%
Male Sterile
msl 6 90%
Defective de16 4 25%
Endosperm
From this list there are three point mutations of utility in the commercial production of hybrids, namely, rust resistance, pigmy and male sterile. The pigmy plants are of quite different phenotype (narrow leaf and other known characteristics) from the dwarf mutants discussed in Example Six(b). However, they could be utilized for a similar purpose, to produce smaller sized hybrids and provide higher plant densities with higher yields. The utility of male sterile plants was discussed in Example Six(B) as well.
In addition to the mutations listed in Table XIII, two important phenotypic alterations were observed which continue to be expressed into a T5 generation currently under study. One new trait involves a line with a maturity which is 12-14 days earlier than the untreated controls. The second is a "broad leaf" expression with leaf widths on the treated lines over 40% greater than those on the untreated controls. The useful nature of the broad leaf characteristic lies in the ability of the plant to receive and utilize more radiant energy per unit time during photosynthetic activity. The result is plant with a more efficient and higher biomass output.
A number of treated series from both the early and broad leaf lines were used in hybrid crossing studies. These plants were selected for either enhanced growth or for altered plant size. When the female line was the treated series, a number of statistically significant yield increases and early maturing lines were observed in the replicated field tests.
This example and previous examples together demonstrate that different donor materials produce significantly different mutations and phenotypic growth responses, as may be seen by comparing the point mutations in Example Six(B) (resulting from soybean root nodule donor material) with those in this example, employing Symplocarpus foetidus as the donor material. New germ plasm is constantly of importance in commercial plant breeding programs and Examples Six(B) and Seven(B) illustrate that although the advantageous expression of a mutation, such as male sterile, may produce similar results, the fact that different alleles are involved in the two examples means that the characteristics in the germ plasm would be expressed quite differently in hybird usage.
CONTROLS
In order to insure that the results obtained in these examples resulted from the combination of subjecting the acceptor species plants to electrophoretic conditions and to whole cells and associated materials of a second species of plant, controls were conducted in which the materials of the second species of plant were replaced by distilled water or by a macerate of plants of the same species as the first species. Additionally, seedlings of the same species had root portions excised and joined together, as well as being exposed to an electropotential difference only. In all cases, no statistically significant difference was seen between any of the plants so treated and untreated control plants. Thus, the results obtained in the examples described in this application necessarily resulted from the inclusion of a donor material from a second, different species of plants.
DISCUSSION
The data obtained in the above examples leads to the inescapable conclusion that the frequency of plant mutations can be increased by exposing plants in their germinal phase simultaneously to electrophoretic conditions and to the whole cells and associated materials of a second species of plant. The fat that some of the mutated plants obtained possess characteristics which appear to be characteristics associated with the donor material of the second species of plant suggests that some genetically associated cell tissue components or macromolecular complexes from the donor species of plant are transferred to or transduced into the intact living cells of the acceptor species of plant, in such a manner as to alter the genotype and/or phenotype of the acceptor, to allowing such altered genetic and phenotypic characteristics be transmitted to successive generations as point mutations or as cytoplasmic transmitted traits. The subsequent discussion and examples supporting such a theory should be taken as evidence of the theory; however, the theory of transduction of genetic materials is not in and of itself essential to an understanding of or a practice of the methods of the present invention. Those methods have been demonstrated by the preceeding examples to be useful in producing an increased number of mutants in a plant population, without regard to whether the instant explanation of how such mutations occur is correct. The fact that the mutations occur is sufficient support for the invention.
The theory as to how the present invention operates is straightforward. It is believed that in the present invention the application of electrophoretic conditions to the cells of an intact organism or whole plant allows the tranduction of genetically associated cell tissue components and macromolecular complexes from the donor species material to the recipient plant species. Migration of these materials would be induced by transmembrane ionophoretic currents, either arising from the natural difference in membrane potentials between cells of different species, or from an externally applied current. The theoretical feasibility of electrophoesis occurring laterally or along, but not through, cell membranes, has been discussed by Jaffe, Nature, 265: 600-602 (1977), and was demonstrated experimentally within the cell membrane and wall by Woodruff and Telfer, Nature, 286: 84-86 (1980). However, as opposed to the present invention, this ion migration was observed and performed by the injection of fluorescent trace-proteins through the cell membranes of an insect ooctye, where they were observed to migrate laterally along "intercellular bridges" or openings, but was not transferred through the membrane barriers without breach of them. Quite simply, the advantage of the present invention is the fact that it is conducted with normal, intact cells of the acceptor species, and at worst with tissue macerates of the donor species. The need to breach or remove the cell wall encountered in all previous techniques is avoided.
Electrophoresis can alter cell plasmalemma permability. This permability is changed by altering the size or current of charge carrier proteins and micropores in the plasmalemma and nuclear envelope. And, as demonstrated by the subsequent examples, it is also clear that the application of electrophoretic conditions allows the ready passage of nongenetic materials through the normal, intact cell wall. Additionally, routine commercial gel electrophoresis techniques demonstrate that some sort of genetic alteration is associated with the method of the present invention. The transmission of certain enzymes, mRNA or tRNA from the cells of the donor plant species to the cytoplasm or nucleoplasm of the cells of the acceptor species of plant alter the rate or path of one or more specific biosynthetic pathways in the acceptor species, which would then alter the phenotype of the cells of the plant. A model of such alteration is shown in FIGS. 10A and 10B, and is described further below.
An examination of electrophoretic technology as a testing procedure was conducted in 20 transduced corn lines and five untreated control lines, from which the test lines were derived. All lines were from the T4 generation of field testing. Gel electrophoresis indicated the presence of 10 transduced lines, or 50% of the total test group, having altered gene alleles. All five inbred controls displayed uniform, unchanged electrophoretic patterns. A total of 8 enzymes, out of the 37 known loci in corn, were examined and provided confirmatory evidence of polymorphism or new gene alleles. In a second test series, 12 enzymes were examined. The test group consisted of 42 transduced lines, 21 from each; of two different in-bred host or control lines. The material from the T4 generation again possessed a high percentage, about 28 percent, of transduced lines having altered alleles, with essentially unaltered or homozygous patterns in the untreated control samples.
Applicant has observed that induced dielectrophoretic properties or long range dipole interactions of a donor material can influence the spatial configuration of organelles within the acceptor cells located within the tissue regions of tranduction. For example, when donors are employed which have a strong, positive dipole charge, that is, a dipole moment much higher than that of water, or donors are employed that have been oxidized and thus receive a net positive charge, those donors migrate from the anode region and pass through the plasmalemma, and associated with the cell nucleus, forming a non-uniform electric field having a maximum intensity at the nuclear membrane. This results in an increase in the frequency of the collection of chloroplasts and other cell organelles in distinct proximity with the surface of the nucleus. In normal, untreated tissue, the nuclear-organelle clustering is observed at a low frequency of perhaps 1%-5% of cells, while in transduced tissue, the frequency in limited regions around the electrode contract zone is observed to be as high as 80%-90% of the cells. Chloroplasts and other organelles are clearly attracted to the nuclear membrane by long range dipole interactions.
Applicant has also observed that the chloroplasts and organelles clustering around the nucleus is not a unique property of one specific donor material. For example, other less dipole substances such as distilled water, when used as a donor, do not produce the nuclear-organelle clustering. In the case of a donor which enters the free space (apoplast) of the host tissue and has a marked dipole moment, but is inert with respect to passing through the plasmalemma, the influence on the spatial patterns of chloroplasts is quite different. In such a case the chloroplast and organelle clustering around the nucleus is not observed, but rather the collection of the donor material in the free space of the cells causes a mass migration of the organelles to the cell wall, the direction depending upon the charge characteristics of the donor material. These changes in configural associations caused by electrophoretic conditions greatly increases the probability level for the exchange of genetic information between the nuclear and cytoplasmic DNA, since the organelles are disposed in proximity with the nucleus. The cooperative, long range dipolar effects occur inside the cell through the microdielectrophoretic interactions between the cell organelles. The existence of such dipole interactions has been postulated by Pohl, Bioelectrochemistry, Plenem Press, New York (1980). By using a ferroelectric material, specifically, barium titanate, Pohl was able to demonstrate dipolar attraction on the outside surface of animal cells. However, as far as the Applicant is aware, the instant observations are the first time that microdielectrophoresis has been observed inside living cells.
Applicant believes the following mechanism may be an appropriate explanation for the observed migratory phenomenon. It is well known that the plant cell wall contains polysaccharides which act as growth and development regulators and chemical messengers. As noted by Albershime and Darvill, Scientific American, September, 1985, page 58, these regulatory molecules are released from the cell wall by enzymes. Different enzymes release different oligosaccharides (small polysaccharides). In a transduction from a new donor species, a donor enzyme complex enters the cell wall matrix and triggers the release of a quite different array of oligosaccharides which, after entering the cytoplasm, redirect patterns of development and form different genotypic associations with either the cell nucleus or the cytoplasmic organelles. As microdielectrophoresis takes place as described above, both nuclear and cytoplasmic interactions occur. This redirection of growth regulators from the cell walls could not occur in the recombinant DNA or protoplast fusion technologies, since the cell wall is necessarily removed in the early stages of the techniques. Further, the enzymes of one plant species may act as isoenzymes of the second plant species and possibly alter the morphogenic properties of the cell. Indeed, there may be enough of a potential difference between the cells of a difference species to facilitate the formation of intercellular cytoplasmic bridges which may allow certain cytoplasmic extranuclear DNA or cell organelles to be transferred from one species of plant to another. The transferred cytoplasmic extranuclear DNA and organelle systems would also exert some influence over the morphogenic determinative components, thereby transforming the phenotype of the tissues.
Additionally, phygocytosis may occur and invaginate certain cell organelles through the cell plasmalemma and into the cytoplasm. Because the cell organelles and cytoplasmic extranuclear DNA synthesize at least some proteins and other materials, which are vital to cell function, the addition of cytoplasmic extranuclear DNA and cell organelles from a different species of pant may cause the creation of enzymes and proteins which are similar enough to the transformed cells' natural products to be utilized by the transformed cell but may, in the process, act as isoenzymes and "isoproteins" which cause the plant to exhibit different phenotypic characteristics, which may then be transmitted to successive generations in a non-Mendelian fashion. For example, in the technique described in conjunction with FIG. 3, the maceration of the donor tissue in distilled water liberates proteins and enzymes inside the cytoplasm of the donor cells, and this can facilitate the transfer of these constituents, because such constituents need pass only from the medium through the cell wall and plasmalemma of the host into the cytoplasm, rather than having to pass through at least two whole cell walls and plasmalemma, as would be the case for non-macerated donors.
The procedures of this invention are believed to involve transductions within the somatic tissues of the host material. The complete expression of a new mutation or phenotypic alteration is not usually observed until at least the T2 generation. For this reason any explanation of what takes place in the host plant after the application of any of the described procedures cannot be based on the concept of a direct, abrupt uptake of donor DNA into the host plant cells during the initial transduction process. The establishing of a fixed genetic expression arising from a transduction appears to be a very gradual process and is believed to occur in a series of stages during the entire cycle of plant development.
For the gradual incorporation of a new genotypic or phenotypic expression into the host plant, the transductions are assumed to be operating within specific biofeedback control systems involved in the plant morphogenesis. To convey this proposed concept of perturbations induced by the genetic transduction process, the least complex of known homeostasis pathways is adopted as a model, B. C. Goodwin, Temporal Organization in Cells, Academic Press, New York (1963). In this simple pathway the alteration takes place at a single active gener locus G@0, which mormally leads to the synthesis of a cellular metabolite m@0 (or enzyme according to the scheme shown in FIG. 10A. In this model, m@0 acts as a repressor or co-repressor at the gene site G@0 through the feedback loop. The main concern here is with the control of protein (enyzme) synthesis Y@0, which regulates the final production of the cellular metabolite.
The assumption is made that the level of the metabolite m@0 is perturbed by the introduction into the cell of a homologous metabolite from a different plant species by means of the transduction process. This new metabolite m@x acts at the cellular locus and augments the concentration of m@0 so that the new level is at the concentration m@1 (FIG. 10B) after the transduction is completed. The rate at which the effect of the transduction m@x is annulled is, for a small perturbation, proportional to the magnitude of the disturbance. From first order chemical kinetics, the level as a function of time t after the transduction is
m@x =a(e@-kt)
where a and k are constants. A very important point here is that m@0 and m@x must be homologous proteins and very similar in their biosynthetic activity in both the donor and host plant systems. if this were not the case, the control loop m@0 and repressor level would be unaffected, or in the case of an incompatible metabolite, the entire loop could be inactivated. This could readily explain shy some species are effective as donors and others are not, and why different tissue regions of the same species respond different as donors.
The perturbation of the normal metabolite concentrations m@0 to a new level m@x would, through the feedback control, alter the rate of mRNA synthesis at the gene site G@0, and a new rate of metabolite production would be established in the tissue of the host plant. As the somatic tissues develop, the entire pattern of gene expression during plant morphogenesis is operating at a different level of temporal organization of nucleotides than would be found in the non-transduced system. As this perturbed, transduced tissue differentiates into meristem regions and ultimately into germ plasm, the kinetics of these altered biosynthetic pathways are transcribed as altered gene alleles, with permanent expression being established in the DNA code. During transcription, the mRNA would contain altered codon sites, which in turn would lead to altered protein synthesis as the polypeptide chains are synthesized on the ribosome surface. Thus we have the situation of the induction of new enzymes synthesized in the epigenetic cycle or enzymatic adaption through the introduction of homologue precursors from another plant species (the donor).
The perturbations of biofeedback control mechanisms within more complex co-repressor systems could account for incomplete or partial masking of dominant alleles in the somatic tissue. In the situation where cytoplasmic mutations arise form the transductions, the inherited alterations may be brought about in quite a different manner. In this case the presence of foreign polypeptides from the donor leads to the possibility that such polypeptides become genetic precursors and may be subsequently imported into chloroplasts and mitochondria, A. Cashmore et al., Biotechnology, 3: 803-808, (1895). The plant genome is unstable and capable of generating variability, Science, 224: 1415, due to changes in repeated DNA units which are more common in plants than animals (more than 75% of all DNA sequences fifty base pairs or longer is repetitive DNA). Repeated sequences are especially prone to undergo loss or gain because they can promote the incorrect pairing of chromosomes during meiosis. If there are multiple copies of a gene, one copy may be mutated and lead to a new function, as in the above transduction scheme, while the previous function is maintained by the remaining members or copies of the gene. Such copies have the characteristics of transposable elements, B. Mc Clintock, Science, 226: 946, with the result that some specialized cells undergo gene activation and phenotype changes. Only DNA loss is irreversible, other DNA alterations such as methylation, chromatin structure, protein-DNA interactions and the like being reversible and modifiable. The mechanisms for all embodiments of this invention are thought to be similar to the above recited model.
Thus, under the application of an electric current across tissues from two different species of plants, transmembrane ion migration occurs, with specific enzymes, their precursors mRNA and tRNA, and regulatory polysaccharides being transmitted from a donor species into the cytoplasm of an acceptor species. Current flow across the tissues also effects the electric charges on the cell membranes and greatly alters membrane permeability and ion pathways through the intrinsic proteins within the cell membrane, which control the transfer of ions and large molecules. With in the cell, microdielectrophoresis alters spatial configurations of the organells, resulting in increased probabilities for the transfer of genetic information between the organells and thereby causing increased rates of mutation. The following examples demonstrate the ready degree of ion migration occuring in cells and germinal plants upon the application of electrophoretic conditions.
EXAMPLE VIII
To elucidate the mechanisms occuring at the cellular level, donors were utilized with known ionic charge characteristics and with both inert and biologically active properties. One type of host tissue consisted of the chlorophyll containing stems of Pelargonium maculatum. Stems about 5 centimeters long and 5 to 8 millimeters in diameter were subjected to two to four hours at about 10 to 20 volt potentials and a current density of about 30 microamperes per square centimeter. The negatively charged, red protein pigment from the Amaranth plant was applied as a donor material in the apparatus disclosed in and according to the technique described in conjunction with FIG. 8, with each end of the host stem contacting a pigment-containing electrode. At the cathode end of the test stem the red pigment migrated through the section, leaving a zone of stained tissue extending several millimeters into the stem. At the boundary of this zone of migration, a microscopic examination revealed the stain collecting of the nucleus of the parenchyma cells. At the anode end of the test stem, the pigment was oxidized and because positively charged. As it migrated from the anode end of the stem it gave the host tissue a dark grey color zone extending several millimeters into the stem. At the boundary of this zone, a microscopic examination revealed a clustering or proximal grouping of chloroplasts in the immediate vicinity of the cell nucleus. The oxidized Amaranth was observed to collect on the nucleus, and through long range dipole intractions (microdielectrophoresis) formed a positive electrical field gradient which then attracted the negatively charged chloroplasts to the surface of the nucleus at the locations of maximum field strength.
Confirmation of these dielectrophoretic alterations in spatial configurations of cell organelles was observed when using a powdered form of carbonyl iron having a particle size of one to 10 microns, with a positive electric charge. When transduced into geranium stems, as in the preceeding example, an electrophoretic migration of 1 to 2 centimeters occured at the anode end of the stem. Carbonyl iron is biochemically active and was observed to be transduced into the cell cytoplasm where, as in the case with the oxidized Amaranth pigment, it caused a long range dipolar attraction of the chloroplasts in the cytoplasm. The chloroplasts were found to be more tightly grouped around the cell nucleus than in the case with the cell Amaranth donor. This is explained by the fact that within the same host tissue and under the same conditions of voltage and time, when compared with the Amaranth, the carbonyl iron migrates over two times the distance into the host tissue. This indicates that the carbonyl iron has a higher ionic mobility than the Amaranth pigment.
A donor macerate of Phaseolus multiflora leaves containing macromolocules and proteins with associated charge groups, when electrophoretically transduced into the non-chlorophyll tissue of Zea mays radicles using the same method as above, causes a clustering of cellular plastids and other cell organelles (too minute to identify microscopically) around the cell nuclei in the anode region of the host tissue. At the cathode region the cell nuclei had a smooth outline and the chromatin structure was uniform. Other less ionic donor substances such as distilled water, when transduced in a similar manner, with the host tissue being either the geranium stem tissue or the non-chloroplast radicle tissue of corn, did not induce the observed spatial readjustments in the cell organelles.
Barium ferrite of particle size 1.3 microns and having a net negative charge was then used as a donor for the purpose of examining a biochemically inert substance which enters only the free space (apoplast) of the host tissue. Using the geranium stem as the acceptor and employing exposures as in the preceeding example, the extent of the migration was far less than when using the more biochemically active materials. The dark stained tissue region was only two to three millimeters into the cathode end of the stem section. At the boundary of the migration, the donor particles cause the negatively charged chloroplasts to migrate and cluster at the cell wall opposite the location of the cathode and migrating barium ferrite. The grouping here was of an entirely different spatial patterning then when using donors which enter the cytoplasm of the cell of the host tissue.
This example serves to teach that in the process of electrophoretic transduction as described in this invention, the donor complex can migrate both through the cell free space of the host tissue, as well as through the plasma membrane into the cytoplasm of the cell. Furthermore, the nature and ionic strength of the molecular dipole charges of the donor can significantly alter the natural, more or less random, spatial distribution of cell organelles in the cells of tissues being electrophoretically transduced. Such altered spatial patterns can greatly influence the probability of the exchange of genetic information between the cell nucleus and surrounding organelles, and thus provide one mechanism whereby mutation rates can be significantly increased. This example also teaches that both organic and inorganic molecular species can enter the plant cell and interact with the organelles in a physical and/or biochemical manner. Components from a macromolecular donor complex produced from plant tissue, also enter the cell and are active in the organelle spatial repatterning.
EXAMPLE NINE
Dry seeds of corn (Zea mays) were inserted between the stainless steel electrodes of the apparatus illustrated in FIG. 8. The electrodes were covered with filter paper pads moistened with distilled water. The embroyo end of the seed was placed upward, or opposite the base plate electrode. At a 45 volt direct current potential a sharp, well defined uniform line of black pigment was observed to develop and migrate up the seed if the base plate was anodic or positive, or down the seed if the base plate was cathodic or negative.
Since distilled water has a very low ionic content, charge transport in the seed occurred through the oxidation of the pigment materials (polyphenols) in the test seed. The migration of these oxidation products, as testing indicates, is linear with time. This linear relationship is what would be expected under conditions of electrophoretic migration. An ionic mobility of about 0.54.times.10@-6 centimeters squared per volt per second was observed, a value which is consistent with the rate of movement of large molecules. Microscopic examination revealed the layer of oxidation products to extend laterally through the tests into the outer layers of the endosperm.
This example provides a graphic demonstration of the movement of large, physiologically related molecules through the plant tissues under conditions of an electrical potential as applied in the methods of the present invention.
EXAMPLE TEN
The frequencies of altered enzyme loci producing polymorphism in corn plants in which Sympolocarpus feotidus is the donor are quite different from the frequencies when using the soybean root-nodule extract as the donor. These different allelic responses are exemplified by commercial electrophoresis tests. In 62 transduced lines produced in accordance with the method described in conjunction with FIG. 3, 15 lines were transduced with Symplocarpus feotidus as the donor, and 47 lines with the soybean root-nodule as the donor. Table XIV provides a listing of the number of transduced lines containing a specific enzyme polymorph, as they occurred within the two donor test groups. Only those alleles showing positive polymorphism are included in this listing; those observed to have only a slight variation are excluded. The enzymes listed are those in which polymorphism occurred in at least one transduced line.
TABLE XIV
Number of transduced corn linesshowing polymorphism
Soybean
Enzyme S. foetidus root-nodule
ACP (acid phosphatse)
6 11
PGM (phosphogucomutase)
4 1
MDH (malate dehydrogenase)
1 1
PGD (6-phosphogluconate
1 0
dehydrogenase)
PHI (phosphohexose
3 1
isomerase)
GLU (B-glucosidase)
1 0
The data in Table XIV demonstrates that in the soybean root-nodule lines the majority of the alterations take place at the ACP alleles. In the lines with S. foetidus as the donor, there were fewer lines with altered ACP alleles and far more lines involving other enzymes. The fact that the two enzymes, PGD and GLU revealed polymorphism in the S. foetidus lines (comprising only 24% of the total test series) and not in the root-nodule lines (comprising 76% of the test series) again emphasizes the influence of the donor type on the final genetic response and range of possible polymorphic alterations that might be achieved by using other donor types and combinations.
Whatever the mechanism yielding the mutations observed when the methods of the present invention are employed, the present invention clearly provides methods for increasing the proportion of mutants in plant generations. The method of the present invention are significantly advantageous over the known methods of recombinant DNA and plasmid fusion techniques, for the reasons that the precise genetic structure of the chromosomes mutated need not be elucidated, time and effort need not be wasted in removing the cell walls, and time and effort need not be wasted in attempting to grow whole plants from isolated tissues. Instead, the acceptor plants are whole germinal plants, which after treatment can be grown in any conventional fashion.
Having described my invention, however, many modifications thereto will become apparent to those skilled in the area to which it pertains, without deviation from the spirit of the present invention, as defined by the scope of the appended claims.
Canadian Journal of Botany, 1970
W.C. Levengood
National Research Council Research Press
Redox currents associated with ion mobility
in stems of plants.
In vivo investigations of bioelectric currents associated with growth, tissue metabolism, and water responses in the stems of trees and herbaceous plants are described. This experimentation is based on a new technique which allows the continuous monitoring of variations in bioelectric currents related to intertissue redox conditions. Exploratory studies indicate relationships between variations in the redox current and turgor conditions in plants. A correlation between the bioelectric current and the difference in oxidation potentials between the plant and ground electrodes is discussed.
In field studies, the current was monitored in different species of trees over a 1-year interval. Similarities in current variations occurred in similar species of trees and were examined in relation to temperature, rainfall, and phenological factors. The current level, examined in trees as a function of radial depth, disclosed the maximum output in the cambium layer. In a study of diurnal variations, a temporal shift in a current maximum was noted along the tree bole.