rexresearch.com
Home
Biological Transmutations and Modern Physicsby
Louis Kervran
Maloine S.A. Publisher, Paris (1982)
ISBN 2-224-00831-7[ Transcribed from an unpublished manuscript of the English translation --- a few paragraphs are unreadable in my photocopy of the manuscript, and some sections are missing. ]
Table of Contents
Introduction
Part I ~ Experimental Proofs of the Existence of the Biological Transmutation Phenomenon
Chapter 2 ~ Experiments Proving Biological Transmutation
(1) Historical summary ~ (2) A few experiments made after 1974 ~ (3) Research by J. E. Zundel ~ (4) Study of the variation of calcium in oat seedlings during germination in twice-distilled water ~ (5) Comments on a study by Zundel on the increase in calcium in oats during germination in twice distilled water ~ (6) Comments on some experiments ~ (7) Reservations on some analytical techniquesChapter 3 ~ Additional Information on Physical Phytochemistry During Germination
(1) Summary of the germinating phases of grain cereals ~ (2) Average curve of the increase in calcium of oats after germinationChapter 4 ~ Photosynthesis
(1) Effects of artificial lighting in photosynthesis for the study of transmutation by cereal plants ~ (2) Photosynthesis limited to traditional aspects ~ (3) Photosynthesis cycle of Hatch and SlackChapter 5 ~ The Devils Advocate
Chapter 6 ~ Problems Related to Phosphorus, Complexity of Phytoanalysis
Chapter 7~ How to Correctly Duplicate a Typical Experiment in Biological Transmutations
(1) Study of calcium variation in cultivated oats ~ (2) Biological conditions ~ (3) ConclusionsChapter 8 [ Missing ]~ Mass Spectrometer Analysis
(1) ConclusionPart II ~ Explanation of the Phenomena By Modern Physics, Theoretical Study
Chapter 1 ~ Explanations From The Atomic Particle Theory
Chapter 2 ~ A Few Examples Of Theories Proposed By Physicists
(1) Process developed by a US Army scientific department ~ (2) L. Romanis contribution ~ (3) Theory proposed by A. Dubrov ~ (4) A few interesting points of view expressed by physicists ~ (5) O. Costa de Beauregards theoryChapter 3 ~ Addenda To The Theoretical Study Of Low Energy Interactions Applied To Particular Transmutations
(1) Effective section ~ (2) Isotopic variations ~ (3) Nature operates at a finer level than man ~ (4) Changes in our understanding of some aspects of physics and weakness of this understanding ~ (5) A few arguments against a unified theory for electromagnetic and weak interactions ~ (6) Intermediary vector bosonsAppendix 1 [ Missing ] ~ Geology
Appendix II [ Missing ] ~ An Extrapolation: (1) Crying wolf ~ (2) One must rethink the concept of energy in living matter ~ (3) Non-electromagnetic energies in living matter ~ (4) Parapsychology and hypothalamus ~ (5) Germination procedure for oat seeds with no external addition of calcium
Appendix III [ MIssing ]~ Weinberg Theory Summary: A few definitions
Bibliography [Missing ]
Introduction
The present work consists of two distinctly separate parts.
Firstly, I cite some new experiments, completed since the appearance of my book which was submitted for printing in 1974 and which was published in 1975: Biological Evidence of Low Energy Transmutations (Maloine Publications, Paris, 1975). That book was concerned with a comprehensive view of a phenomenon which calls for precise analysis. This led me to limit research to a single aspect: to show that, however precisely one studies the germination and cultivation of a plant, comparative analysis of the seed and of the whole plant coming from a seed as nearly identical as possible, without the possibility of having an external contribution from some mineral, show that there is, in a very significant way, a "creation of matter", "appearance" of an element, thus an atomic "transmutation", which is confirmed by various methods of analysis utilizing the most sensitive, most specific, and most modern techniques of physics.
I limited myself to studying variations in the amount of calcium without studying what element or elements could have given rise to this variation by this atomic modification. Such a research project could be very complex indeed because there could be several origins as a function of the species of vegetation (or animals, higher or lower, even microbial), confirmed experimentally: the calcium could come from potassium, from magnesium, or from silicon, by either separate or simultaneous reactions. So we have there a completely different point of view. I wished to limit the subject so as to produce irrefutable evidence that there is indeed in life forms a phenomenon which too many people have wished to deny for untenable reasons. I will demonstrate it and limit the study to the variation of calcium in a single cultured species, oats, in order to firmly show that such experiments can be reproduced, that the conclusions result from hundreds of experiments and thousands of analyses and are amply demonstrated. Accordingly, we are concerned here with an objective contribution [apport] and not with a subjective deduction.
Next is shown an example of explanation placing itself in the framework of the most recent atomic theory, that of "neutral currents" already sketched in the last chapter of my work of 1975 cited previously which included a "Terminal Note" of 1974 from the great French physicist of international stature, Oliver Costa de Beauregard, theory confirmed by a specialist in elementary particles, Bernard dEspagnat, director of the Laboratory of Particle Physics of Paris. We are concerned here with a very young branch of nuclear physics which is evolving very rapidly and I cannot even dream of following its most recent discoveries, out-of-date before being printed. I will make a very condensed review of the situation in this science toward the beginning of 1981, keeping in mind that the authors of basic principles of this theory received the Nobel Prize for Physics at the end of 1979. That is to say that this aspect of weak energy interactions is now adopted by International Science. This section seemed indispensable to me because too many physicists, and along with them scientists from various other disciplines, consider transmutations only a phenomenon which recapitulates strong interactions. Blinded by the atomic bomb, they have not thought that there were also low energy transmutations, from which they produce a stubborn and sterile opposition to my work.
Here I give only the current status of a theory that is rapidly evolving. It will probably be superceded in a few years, or more or less revised, but it is necessary to show that it is not rejected by nuclear physics avant-garde, that if I have been correct too soon (for them), nevertheless the transmutation of certain elements by a biological action is in no way mystical; it is an explicable reality in conformance with a theory which is now very classical and official and still ignored by too many scientists. Here they will find an incentive to study more deeply this new entry into particle physics, unfortunately not possible to lay out in detail because new facts are turning up all the time: in 1980 did not one come, does it not seem, to produce evidence that neutrinos (basic particles of weak energy transmutations) have weight when all calculations prior to 1980 were conducted under the hypothesis of a null mass?
I will show, then, that:
(1) transmutations by living matter and with weak energy, do indeed exist; and
(2) that they fall within the framework of classic theory of weak energy interactions.
Part I
Experimental Demonstrations of the Existence of the Phenomenon of Biological Transmutation
"It is absolutely impossible to prove a priori the impossibility of a fact" (Bergson).
General Overview
Let us start out by considering a statement by Claude Bernard which will always be timely: "The experimental method consists in revising theorems and not in preserving them. Theory must adopt to nature but nature need not adapt to theory". Pasteur, on his death bed, confirmed to Renon: "Bernard was correct when he said: 'When one encounters a fact which conflicts with a dominant theory, one must accept the fact and abandon the theory, even though the latter is supported by influential people and widely accepted'.
Experience has taught me just how close to truth was Claude Bernard. To talk about the transmutation of matter in a biological environment seemed to be a risky bet for certain theoreticians. In practice, however, it is certain that this phenomenon had appeared more or less clearly to a number of observers at every intellectual level. Many professionals in various disciplines have expressed this openly. It seems that the irrefutable facts were there known to everyone, but refused acknowledgment by certain persons because of their timidity. Or perhaps, having indeed seen the facts, they still did not dare say so".
Thus, between the two world wars of the 20th century, a Swiss agronomist, Pfeiffer, who emigrated to the USA, called attention to the fact that a gardener perceived that his land lacked calcium when his lawns were covered with daisies (or with buttercups). It is obvious to everyone. The turf is composed primarily of rye-grass, a calcareous plant --- one which consumes calcium --- which is therefore an indispensable component of the soil in order for it to grow well. By contrast, daisies and buttercups, etc., are calcifugous plants: their development is not satisfactory unless the soil is acid, almost completely without calcium. Having had the curiosity to analyze the ashes of the daisies, however, Pfeiffer discovered that these plants, which fled from calcarious soil , were rich in calcium. Could it be that a balance is established, something like a symbiosis between rye-grass and daisy, precisely because the daisy produced in the soil, by developing and dying, the calcium needed for a good growth of rye-grass? He did not push his research any further, simply accepting the assumption that in this soil, from which the calcareous component was sucked up by the rye-grass carried off after having mowed the lawn, there was a creation of calcium by the daisies. That was scarcely orthodox and he did not dare state that position openly --- to protect his career, perhaps?
A discovery of the same order may be credited to horticulturists who were intensively growing heather land flowers for resale. The calcifugous plants (azaleas, etc.) did very well in a "good" heather land for several years. Then, little by little, the culture declined and it was necessary to abandon plantations of this sort or to renovate the arable soil. In horticulture we find eminent engineers and superior technicians who thought that the solution to their disappointments could only be realized by chemical analysis of their land. Accordingly, they discovered that this land, originally acid, had become basic, or occasionally neutral, clearly richer in calcareous substance, even though taking into consideration that they were growing calcifugous plants they had certainly avoided laying on any calcareous fertilizer whatsoever. Where did this undesirable element come from?
Did they visualize an elevating of calcium from the subsoil by capillarity? But certain cultivation, from all evidence, was not on a calcareous subsoil. To get to the bottom of this they removed the stratum of arable soil right up to the subjacent layer of impermeable clay, laid out some sheets of plastic on this layer to insure that there would be no reestablishment of calcareous substance by any sort of unfortunate migration, vertical or horizontal, and then covered the plastic over again with a layer of good heather land soil which had been carefully analyzed.
After a few years there was a repetition of the preceding observations: without bringing in any calcareous substance the soil was enriched in calcium. Having become acquainted with my studies they asked me at a conference to explain to them the mechanism by which I thought: in my opinion there had been production of calcium by the rootlets remaining with each uprooting and accordingly absorbed by osmosis by the roots. What other explanation can there be?
But it does not suffice to deduce and affirm. It must be demonstrated scientifically by systematic and rigorous experiments. I did that. One will find in the present work irrefutable demonstrations that certain calcifugous plants produce clear evidence of calcium in clearly measurable quantities, absolutely without any possibility of error, excluding the possibility of a change in solar radioactivity. Agronomists who know how to observe and whose judgment is not obliterated by obsolete dogma clearly admitted this since it was a phenomenon well known to their profession for some decades, although still formulated with too much timidity.
I should also cite results of analyses given by semi-official Tables of Nutrients, often referred to under the name of Tables of L. Randoin, accounting services of the National Ministry of Education. They do not make precise distinctions for different varieties; they only give averages for genera and species and sometimes the analyses --- without precise methods --- have been borrowed by different authors. Lets take the case of the soybean. It gives the composition of the seed. We give here only the percentage of calcium, the element which will mainly be studied in this book.
It gives: 280 mg per 100 gr of material. In soya sprouts, it indicates for Ca: 48 mg per 100 gr
These values are given without precise description of cultivation and methods of analysis (all chemical at this time). Accordingly, they cannot be compared without some calculations. The seed is given with a percentage of water of 7.5 gr per 100 gr of material, while in the shoots there were 86 gr of water per 100 gr of material. From this fact, proceeding from equivalent dryness (that of the seed being the starting point), the percentage of Ca, which is 48 mg (for 86 gr water) would be 554 mg in lieu of 280 in the seed, for 7.5 gr of water. The increase in the sprouts then, is in the neighborhood of 96%, which is clearly in the order of magnitude of the increase of Ca in germinated oats, as we will see further on.
So then, did the plants only accomplish a chemical exchange? Or did they accomplish alchemy? This word generated fear and was rejected with horror by scientists subjugated by concepts of physics much too recently established to be lightly set aside.
Too many physicists believed devoutly in laws which they considered to be general, universal, and applicable under all circumstances. The evidence of facts which could not be explained by their theories, which could not even come close to disturbing the pattern of their professional commitments.
For them it was actually profane to think that a transmutation could be accomplished by a living organism. This was evidence of feeble-mindedness... Indeed, people were obliged to admit that transmutation phenomena could be produce din nature and in the second third of the 20th century we even succeeded in reproducing radioactive transmutations (1935). At the same time, non-radioactive transmutations had been artificially produced since 1919. Then it was in 1945 that the atomic bomb had a formidable impact on the thinking of physicists, blocking all critical thinking in far too many of them.
As a result of my previous studies, first published in 1936, I was officially appointed at the national level, to follow closely the creation of nuclear physics essentially to promote security measures to prevent biological effects of atomic radiations. For a period of 20 years, up to my retirement, I retained these functions, periodically reconfirmed by council orders. Thus, I was at the interface between physics and biology before the emergence of atomic energy. As a matter of fact, I was present at the birth of nuclear physics and was able to follow all its developments. Because of my official duties no laboratory classified secret was closed to me.
In 1936 the author published the first results of his experimental research showing that the human body does not follow Ohms law, that its resistance varies as an inverse function of voltage applied, which explains values which are not independent of voltage output from the ohmmeters employed (for example, see Outlines of Industrial Medicine by Prof Simonin, Maloine Publ., Paris)
These researches were interrupted by the second world war, and in 1940, the author was arrested for Resistance, incarcerated at Fort Montluc in Lyon, and found guilty without appeal. Having served his time, he participated in setting up the Southeast Resistance. In 1944 he was appointed Prefect by the general assembly of the Committees of Liberation of Savoy, and then given the duties of Regional Prefect of Savoy-Dauphine. He received the Medal of the Resistance. Shortly thereafter he re-entered his original cadre as director in charge of scientific functions at Paris for a period of 20 years.
That is why I assessed the value of what was obtained with certainty in this discipline but also the limits of our knowledge in accordance with the trend of the times. It seemed to me that, primarily in biology, but also in physics, unverified data were being assumed by inference [extrapolation], even though they were contradicted by certain observations which oriented me along channels which were being ignored completely by most atomic physicists who were unable to question their own understanding.
"You should discuss the matter with your peers", they once said to me. But who were my peers? As a function of my duties I was also named "director of conferences" at the University of Paris. That was, in fact, the official designation, as represented in the Deans teaching directories. Accordingly, were my pers these tens of thousands of teachers on our faculties? Could I put myself in the same structure as tens of thousands of professors of higher education when I had a unique function in France, recognized by an official appointment (and even several inter-ministry groups representing all the ministries scientifically interested in atomic energy, by one title or another, also appointed me to represent them in the inter-ministerial commissions which were obliged to take regulatory actions in this domain requiring expertise in both biology and atomic physics (Atomic Population Protection, Public Health, etc.)
Indeed I knew, having been a member of the examining committee at the doctorate level (before the title was degraded by creating the "third level doctorate") how impossible it is to avoid that in a huge corps of tens of thousands of members some diplomas slide by that are really below any standard. If they are, in the overall, about one quarter of mediocre, about one-half average, and one quarter good from which one can select out a true elite, several working together --- I had no practical way of deciding who was a member of this elite group, and, in any case, it was not up to me to make such a necessarily subjective and arbitrary decision. I make no claim to be universal but I would recognize who in the national scene had an "international value" and that my duties enabled me to consult, moreover, if need be, on any detail, in order to achieve a synthesis, which is becoming more and more difficult to realize because of intensified specialization which no longer leaves a place for any but the "analysts", and rejects the "synthesists".
In 1950 I began to publish the first results of my research showing that living matter, both animals and plants, accomplished transmutations of elements. These transmutations were observed in man, animals, microorganisms and plants. They were transmutations which, from all evidence, had nothing in common with high energy transmutations which are the only ones which the majority of atomic physicists are inclined to accept and the only interactions which scientists in general accept without any reservations. However, this did not block official acknowledgment of the value of my work and in 1964 I received recognition with a Legion of Honor ribbon.
I has experimentally produced irrefutable evidence of the existence of facts which could not be explained solely in terms of "chemical" biology. I had evidence of phenomena of non-radioactive atomic physics which could not be explained by classical atomic physics of this time. But I had established that the only thing which could account for the observed results was a nuclear physics which remained to be more precisely developed and which I was the first to express in clearly stated formulae of nuclear reactions.
(1) "Life Is Nothing But Chemistry" ~
To a greater and greater extent, throughout the 19th century and then more fully in the 20th century we have been taught that all biological phenomena depend upon chemical reactions. I certainly do not deny the truth of this obvious situation. But that is only part of the truth. If one desires to reduce everything to chemistry one is led into serious errors with respect to human, animal and vegetal biology.
In advanced agricultural schools and faculties of science and medicine one still sees it advanced with laughable self-confidence that, as an example, water is always water; there is only one formula for water: H2O (which should be expressed less rigidly, for we can have H3O4, etc, and the diversity of snow crystals shows that... but I do not wish to deal with chemistry). Take some grapes which have been slowly dried out, then soak them in pure water which bears a trace of mineral or organic matter. Everyone knows that grapes thus reconstituted do not taste the same as and have other properties than fresh grapes. Likewise, chemical analysis shows that composition with respect to carbohydrates, lipids and proteins differs between fresh and dried grapes and that these differences are not simply a matter of water evaporation. All nutritionists who are not blinded by dogma recognize this and take it into account.
Otherwise stated, simplified chemistry fails to account for modifications of molecular structure resulting from a physical phenomenon such as gradual evaporation. But that is nothing more than an aspect which can be easily explained, for example, by classic procedures of stereochemistry. In my publications and in conferences I have called attention to the transformation that the organism forces on carbohydrates to generate lipids (one can fatter a pig on nothing but potatoes which are rich in carbohydrates but poor in lipids). In a diet nothing will suppress lipids for people who have a tendency to fatten on carbohydrates. It appears that their organism provokes this transformation into lipids as a result of some metabolic disturbance which may have a number of glandular or alimentary causes such as a magnesium deficit, etc. One does not cure the effects by forgetting the cause. And this is true throughout biology. We should remember campaigns against food substances rich in cholesterol, a normal product of physiological catabolism, If they are not provided in the diet the organism will, nevertheless, fabricate them. The unbalance is to be discovered in the process of elimination, not in the process of absorption. It is not without interest to recall that if certain individuals get fat on white bread it is because this bread is poor in magnesium due to a sifting procedure which is much too gross, all the "rich" part of the grain having been eliminated to be resold separately at very high prices. But few simple formulae have survived in dietetics, alas.
And, likewise, how many simple formulae have survived with physiologists? For the majority of them --- and the guilty are teachers, for the most part, particularly chemists --- a carbohydrate is a tertiary composition where the hydrogen, oxygen and carbon are clearly defied constituents. For example, for decades they have accepted without reservation that pure saccharose was always saccharose, that there was one possible formula, confirmed by all methods of chemical analysis. Accordingly, its biochemical properties are always the same. But this is not true and if the chemical formula remains clearly the same, nevertheless its biological properties differ in accordance with the origin of these saccharose substances. Thus we can distinguish a saccharose from a beet, which does not have the same isotopic composition as can sugar. Nature makes use of several methods to separate isotopes, to modify the isotopic composition, and in critical proportions, making a lie out of the simplistic conviction of too many physicists for whom the isotopic composition of an element is obviously constant. In the bibliography at the end of this publication one may refer to the works of Bricout referring to various authors. We see that it is now a common practice for the Service des Fraudes and customs office to use mass spectrometry to reveal if a saccharose comes from beets or sugar cane, because the import quotas differ as do the prices.
The confusion has persisted for a long time because, still too often, classic instruction only takes into account the Calvin cycle in considering the function of chlorophyll. But this is the cycle utilized by the beet and most dicots and too many publications still ignore the cycle of Hatch and Slack, used in photosynthesis of sugar cane and the majority of monocots. They have understood better that by adding tap water to dried fruit or to concentrated fruit juice one cannot obtain the qualities of fresh fruit and fresh juice. The water in a fresh fruit is not the water found in rain, in an irrigation ditch, or drawn from the soil; it is a different compound at the subatomic, neutronic level. The number of neutrons differs as a function of the origin of water but the number of protons and, accordingly the number of electrons remains the same and it is therefore impossible for chemistry to differentiate them. I will return to this later on because it has various implications which many chemists, atomic physicists, agriculturalists and nutritionists have not taken into consideration. This situation will be considered more carefully with respect to the functioning of chlorophyll, a mechanism which is fundamental to this differentiation in isotopic behavior. However, I will not undertake a detailed study of that because the isotopic separation provoked by the metabolism of the plant is not a transmutation. In a way it is a kinematic operation between heavy and lighter isotopes. Heavy hydrogen, or deuterium, is approximately twice as heavy as ordinary hydrogen which has no neutron as opposed to one neutron in deuterium. The speed of the reactions has a substantial effect and Ponticorvo, in a thesis written in 1958, cited by Bricout (cf. reference section) calls attention to a reaction which leads to a 70% diminution of deuterium which, from all evidence, is very significant and cannot go unnoticed in a spectrometer.
This property of living matter is often not taken into consideration by physicists who undertake analyses with an a priori assumption of isotopic constancy or, on the other hand, by biologists who use radioactive tracers and generalize conclusions from data which are only observed under very limited circumstances. A more precise study is definitely called for, particularly in view of the fact that a radioactive isotope destroys cellules and thus opens a pathway which modifies the behavior of a stable isotope. I dont mean to say that we should abandon using radioactive tracers, but we would be well advised to be cautious in making judgments based on their repeated use. Belief in constancy of isotopic composition is an idea too easily adopted without a critical attitude and it sometimes leads to errors.
(2) Lavoisiers Law ~
The preceding comments show why it is wise to be cautious about general application of Lavoisiers laws in biology. In no way do I reject these chemical laws, but let us leave them in their place. The statement that "Nothing is lost, nothing is created" actually is just a play on words creating a certain impression, Reality is more complex. Chemistry is indispensable in order to understand molecular transformations, which were the only sort of transformation which could be demonstrated in Lavoisiers time; but at the atomic level they become inadequate even for those who take the position that in living matter there is nothing but chemistry, since phenomena occur there which can only be studied in terms of physics; electrical effects, pressure, heat, movements, etc. This is true even if one takes the position that chemistry is simply a branch of physics and is explained in terms of displacement of electrons. The present study will show that living matter employs energies which are not electromagnetic, that nature also operates right into the heart of the atomic nucleus, which has nothing to do with Lavoisiers laws. Lavoisier was completely ignorant of this aspect of living matter, even though his contemporary, the great French scholar Vauquelin, suspected it. However, he was a century in advance of physics and had no practical way to study the phenomenon.
Even in our time there still exist pseudo-scientists (by profession) who take the position: I do not doubt your analyses, but if you no longer find an element, that is because it has gone off somewhere else where you have not searched for it. Because, for them, nothing is lost.. certainly, I could not analyze out an entire human body at one fell swoop. Studies made on calves were subject to the same criticism: the analyses could not have been carried out on the entire calf. And so I worked on lobsters, but in this case the reproof was that I had only used 8 animals. I carried out my studies on 48 mice and for various reasons the animals were totally dissolved in acid. This is a complex operation because one risks the formation of soaps, by saponification of fats; there are difficulties in dissolution of the keratin of the hair, etc. I have described these various experiment sin previous publications. The complexity of the procedures gave too much opportunity for criticism for there is nobody more deaf than he who does not wish to hear. And so I decided to proceed only with relatively simple operations, working only on plants and finally limiting myself to the study of variation of one single element, calcium, in a single plant species, oats, under hydroponic culture, without involvement of a complex culture medium, using only synthetic water (hydrogen and oxygen) or double distilled water. Water demineralized on ion exchangers was not always sufficiently pure. Or it might be necessary to distill permuted water.
In this process I found myself in cooperation with Zundel and his studies will be cited further along.
Working with tens of thousands of seeds I could now talk only in statistical terms, in terms of averages, where any individual difference between seeds more or less vigorously disappeared before the law of large numbers. These analyses, applying to hundreds of cases, lead to proof that one finds, in a calcifugous plant, a great deal more calcium in the plant than there is in the seed from which it came. By incomprehensible quibbling certain chemists have extracted from this only a proof that they were incapable of making a good analysis. For them the discrepancies in calcium composition derived from the fact that in the seed there were undetectable "hidden forms" which became identifiable after germination, after mineralization by oxidation. Now the "organic" form in the seed could not be discovered by titration. Whence, evidently, an augmentation of Ca in the plant. Now other chemists equally incompetent, equally dogmatic, explained the contrary, taking the position that in germination insoluble forms were produced, which could not be measured by titration, and that most "seriously" of all. What a pity for their students.
Now variations have been discovered not only with respect to augmentations. For certain elements in a plant species, in extra pure water, with acid pH, there can be reductions. I will take the opportunity to deal precisely with these aspects of analytic methods.
Certain people suggested to me that I call this book my Testament", considering my present age. But, in science, I take the position that there is no such thing as a "last will". I would be presumptuous of me to think for one instant that anybodys scientific contribution can be regarded as advice to be followed in the future. Science itself is in full charge. In no way can we anticipate when the results of scientific progress will be replaced, when we will have attained the asymptote of the curve of intellectual growth. Thats why, when in 1979 I thought to write the present publication, I could have entitled it "Twenty Years Later", a title which would scarcely relate to me! It would more appropriately be my "swan song".
More modestly, then, you will find in the following pages a tally of more than 20 years of work done by different researchers who became interested in my first publications showing that there were in the metabolism of living matter, both animal and vegetable, some aberrant phenomena which could not be explained simply in chemical terms. Only through nuclear physics could we come to understand such findings resulting from irrefutable investigations and I used to advance the statement: there are, in that which lives, certain transmutations, which, to abbreviate and to avoid confusion, I called "biological transmutations", avoiding use of such expressions as fusion and fission, which called to mind atomic bombs, since we were certainly concerned here with a phenomenon of nuclear physics totally different from high energy interactions, which are the only ones which the majority of classical physicists were studying.
I explained this in my first article, which appeared in 1960, and even more fully in my first book, Biological Transmutations, which was issued by Maloine in Paris (1962). This publication at the end of the same year, was translated and published in Japan, at the instigation of G. Ohsawa, then some ten other publications followed in France and abroad with so many reprintings that, in 1980, somewhat more than 100,000 copies had been issued. The "biological transmutations" had, accordingly, made their mark despite certain oppositions which are inevitable when one upsets accepted ideas, taught traditionally, sacrosanct, and distributed via all channels: books, reviews, television, radio, where certain people had established their position such that they could no longer recognize their error, which is not scientific. But I received numerous positive supports from eminent scientists, without loss of professional position, above and beyond dogmatism, who determined that my studies were unimpeachable, and I was able to accomplish numerous publications, conferences, etc., in France and abroad in order to subject my material to discussion. For we had recognized and accepted data. It remained to explain the phenomenon; and any explanation is more or less subjective and dependent upon knowledge which is more or less accepted at the time.
In any case, after 20 years, Science has evolved. Thats why the present publication seemed to me to be necessary. I do not intend to disavow my first publications, but rather to clarify, to abstract certain prospective notions which have not been realized, which will not arrive, perhaps, until later on. It is also intended to set aside certain over-extended representations destined to concretize a phenomenon produced in our cell structure at an imperceptible level. To explain is to attempt to force facts into the framework of theories which the majority of scientists accept at a particular time. But these theories have evolved in 20 years. Accordingly there are representations to be reconsidered and it is not excluded that we may definitely throw out some comparisons which I made about 1960 and shortly thereafter. I will show as an example that very recent studies recall certain representations which I proposed some time ago.
Certainly, the facts and the experiments remain and retain their value for all time, except for experimental error which was not detectable at the time. It is the explanation of these facts which has evolved.
That led me not to follow through on certain reprintings. For, in 1974, a sharp turning point in theoretical nuclear physics led me to rethink the entire theoretical portion of my studies and that will be seen in greater detail in the second section. For this reason I ask the readers of the present publication to disregard explanations of my books in review articles or sound recordings, etc., prior to 1974. In a sense, they no longer have anything but historic interest. But I do not totally discard certain analogies which can be useful and are not all absolutely false. Certain recent advances in exploration of the atom show that some notions that I expressed in the early 1960s seem to be confirmed but cannot be generalized in a simplistic manner. This infinitely small world escapes out senses in a way which renders it unwise to bend it into a set of images structured for our senses on a multi-molecular scale. Only my book Preuves en Biologie de Transmutations a Faible Energie (first edition 1975) will be reprinted. That publication contains a general perspective of the principal experiments which enabled me to conclude that there are effective transmutations of elements by biological mechanisms and what are the extensive applications that they have found in medicine, in agriculture, and in dietetics. The present work scarcely touches on applications, constantly changing, and attempts primarily to make available to scientists the irrefutable indications showing that such transmutations do certainly exist and that we now have an explanation in theoretical physics which permits their accommodation in classical studies. Therefore this is a complement to the 1975 publication which will be kept up to date in future printings.
I wanted the present book to adapt to meet the most up-to-date theories of nuclear physics and also to be an inventory of various researches specially undertaken and irrefutable in terms of scientific method. I wish to set forth certain material involving details which must be carefully considered because experience has shown me that well-known specialists were not capable of obtaining transmutations placed within the framework of theories which were unfamiliar to them. Due to some sort of professional defiance or to ignorance resulting from over-specialization, but in any case, unfortunately, most difficult to get rid of, they cannot see the decisive role of this or that biological situation. Routinely they are led into errors because they wish to transpose the concepts they hold to a new science which demands that they set up certain practices and theories which are completely out of line with those that they have always employed.
Not everyone can set aside the concepts into which they have been formed (or deformed) ever since their earliest schooling. I will sow a few examples, but it is obviously impossible to address every aspect of this new problem which leads to calling in question an array of notions which are regarded as classic. One must reflect shift ones wisdom, avoid automatic reaction. For all of us, when we do not understand we must wait for the moment when we will understand.
The problem studied here remains in conformity with known laws of physics. The reason that we have studied transmutations for the most part among biological channels for the past 20 years is because this milieu gives a relatively easy means of producing them and then reproducing them. And this is accomplished under very precise conditions which preclude generalizing in a naïve and infantile manner. The energy action which precipitates the transmutation demands a combination of certain specific conditions; it does not appear in a seed kept dry but does show up after a few days in a seed placed in a condition permitting germination. There is then produced a synthesis of enzymes which modify the spatial structure, the stereochemistry of certain proteins constituting ADN, ATP, etc. But this structural modification is just a preliminary stage. It leads to exponential multiplication of the capture cross-section (effective capture cross section) of molecules with cosmic neutrinos in this ocean of particles in which we bathe.
We are not dealing here with some mysterious property which calls in some sort of vital principle more or less well formulated from another point of view. I have, in fact, been able to show that certain minerals, in combinations in metamorphic rocks, so-called because experimentation and observation have shown that they can change their form: these minerals can show transmutations in line with the same theories --- however, on a different scale, because stereochemical modifications which occurred with the application of temperature and pressure which "fluidifies" the mineral makes atom displacement easier and these rocks, having come under the influence of cosmic neutrinos, modify their atomic composition both qualitatively and quantitatively. In my 1975 book I dealt with research done under a pressure of 50 kilobars and a temperature at the 850° C level. And so we also have applications in the study of mineralogy and I will come back to that briefly, from another point of view, because eminent geologists have been able to advance explanations of phenomena which were completely incomprehensible in terms of classical theories.
Chapter 2Experiments Establishing With Certainty Certain Biological Transmutations
(1) Condensed History ~
I refer the reader, for more details, to my basic book of 1975: Proof in Biology of Weak Energy Transmutations (Maloine, Paris). None of my prior publications will be printed again in full.
In the title of this book I did not retain the expression "biological transmutations" and replaced it, as in other publications, after 1963, by the more general expression "weak energy transmutations" because I was fully convinced, since my first publications, that this phenomenon demonstrated by numerous experiments was more general than simply biological and I referred to it in my second publication of 1963 by the term "Natural Transmutations". In the present volume, in order that one not lose sight of this general aspect, I have summarized in one chapter a few applications in geology, but it is a very limited presentation of several studies appearing since 1975. However, it is not my intention to take applications into account here, and it would be good to have a book expressly aimed at studying the impact of weak energy physics specifically on mineralogy to get a view of everything available in this domain. I will refer to several publications, one of which has more than 90 tightly written large pages, and there are some publications which have appeared abroad. It would certainly be desirable to distribute a synthesis of the essential experimental studies along these lines because one must have very costly materials for this sort of investigation while in biology any laboratory can do research without great cost.
However, here I wish to convince people that transmutations of elements can take place with low energy in living matter under conditions which will be made precise for one must never say that a phenomenon is general, that it occurs everywhere all the time. What I wish to show is that my researches are a consequence of putting to work weak energy interactions, and not a consequence of high energy interactions which have been the only sort of interactions that most physicists have considered since 1974. I save the study of physics for the second part. This is fundamental because too many scientists have their minds twisted by physicists subjugated by the atomic bomb which led too many of them for some 30 years (a whole generation unfortunately prolonged in distortion by those who continued teaching) to deny the existence of and fail to see the possibility of weak energy transmutations. However, the majority of truly great physicists did not lose sight of natural weak energy transmutations, and I will come back to that, but these weak energy transmutations are a phenomenon which is no less striking than atomic explosions and the contribution of these truly great physicists was more modest than that of the majority.
When I discovered these biological transmutations and decided to publish my conclusions, I was not thinking of integrating these findings with what was known (actually very little) concerning weak energy interactions. More to the point, I did not know that very early experiments, conducted over a century and a half, had demonstrated the creation of certain elements and the disappearance of other elements (or, more precisely, the augmentation of some and diminution of others). Biological observations in animals and plants were numerous and varied, but for very "humane" reasons they had been kept under the blanket and were relegated to trivial publications which people were reluctant to quote and therefore they remained practically unnoticed.
But millions of people, via my publications, widely disseminated reviews, by radio and by television (and because my official functions made silence impossible) learned about my studies and among them were those who were aware of previous experiments and several called these previous experiments to my attention. I should recall, for example, that the major popular science review Science et Vie (Science and Life) devoted several articles to this from 1960 to 1963 in some 350,000 copies. Europe No. 1 (June 1961) distributed my 40-minute interview with Jacques Mousseau and, previously, the Belgian television 819 devoted about a quarter of an hour to these matters in December 1960.
If the above mentioned publications remained in the dark for the most part it was because they were premature in the sense that they were incomprehensible and too many people deny that which is not understood even though the facts are indisputable. However, I advanced an explanation by a mechanism which had nothing in it which was mysterious for the 20th century because it was in some ways parallel to fusions and fissions of the new atomic physics born with the century.
Lacking an accepted scientific explanation these publications were often rejected out of hand on the ground that they resulted from experimental error. Furthermore, it was impossible because it would be a return to alchemy. That had been definitively thrown out by science of the 19th century. One could no longer go backwards. It was absolutely necessary, however, to proceed to the evidence of the turn of the 20th century.
The discovery of radioactivity demonstrated in a striking manner that transmutation of elements was impossible to deny. One could study it better when, in 1919, the first forced transmutation was achieved while in 1935, there was a successful artificial production of new radioactive substances.
In 1963, the atomic physics professor of the Conservatoire National des Arts et Metiers made available to me photocopies of several dozens of pages in which Freundler, a Sorbonne professor, condensed, in a 1928 book, studies conducted for more than 10 years on the production of iodine by algae. He is the first, to my knowledge, who saw that there was a connection between the tin of the granite support and the iodine in these plants. He had sensed the type of reaction that I indicated but he had not been able to convince anyone of this. He had come too soon and his calculations had a weak point. The balance of charges and masses was defective because the neutron was unknown at that time, not having been discovered until 1932. But nobody else, even after 1932, dreamed of reconsidering the problem which was nevertheless cross-checked, as it were, by converging studies. I touched on the work of Freundler to a certain extent in my book of 1963 on natural transmutations, which, after two editions, was not printed again.
Readers of books failed to see a good many lines which were rediscovered and took on new dimensions when those readers saw my first publications. It was in just this way that a friend of mine called my attention to a passage of Flaubert in Bouvard et Pecuchet, a publication which challenged the science of the 1880 era in a series of critical dialogs. One chapter was written by Flaubert under the inspiration of Regnault, a physicist well known to schoolboys because of his Thermodynamic Tables, specific heat, etc., and by Giraudin, an agronomist of world-wide reputation. Flaubert emphasized that the great French chemist Vauquelin (a contemporary of Lavoisier but more open-minded than the latter) had demonstrated that a chicken fed exclusively on oats laid in its eggs and in its droppings more than four times as much "lime" as it had ingested with the oats which had been analyzed beforehand. He provided a balance sheet for the "lime", which was what we would today call calcium carbonate, and also that for "lime phosphate" because the balance sheet for phosphorus was also modified. But the spectacular finding was the augmentation of calcium. There was, according to Vauquelin, a creation of matter. His memoire had been published 19 January 1799. He tried to see what could have been reduced in order to give all this "lime". But in those times people did not know about the atom and he did not conceive of certain possible origins. I was able to obtain the original of this remarkable piece of work and discovered that they knew very well how to make precise analyses of calcium at that time. But Vauquelin knew how to isolate and discover certain "simple substances" and he was a very talented and clever experimenter. Furthermore, in those days it was the main boss himself who made the analyses and not a laboratory assistant. In my 1975 book I devoted 10 pages to presenting this remarkable study which, in my opinion, constituted the oldest and one of the most serious experiments before the atomic era on biological transmutations, an expression which had not yet been formulated in the 19ht century and was much less available in the 18th century. I did not mention this test in my first publication because I did not discover it until about 10 years later.
That indicates that Lavoisier had produced an absolute law which was true within the framework of chemistry but in chemistry only. Vauquelin showed that there was something else in living matter and that the problem is more complex than in the chemistry of non-living material. This shows the great historical importance of this 1799 publication.
I also refer to my 1975 book for the very important studies published from 1875 to 1883 in Germany by Von Herzeele. He did numerous experiments on a great many species of seed germinated in a dust-free situation with distilled water containing a mixture of two salts. One of the salts always contained a constant anion and a cation which varied with each experiment. In other experiments this was reversed: constant cation, variable anion. We see that this investigator anticipated the phenomenon of transmutation and was studying the correlation between the augmentation of one element and the diminution of another. But at that time atomic structures were not known. He was then about 20 years before his time. However, the results that he obtained are very important since, after Vaquelin, we have a second stage, conducted scientifically, a valuable example of researches on variations of certain elements as a function of metabolism in the germination and growth of various plants. Certainly, Von Herzeele (and several others before him, not knowing the structure of atoms, completed certain experiments which had no significance, but some of his experiments are valid. In addition, being a chemist, that certain reactions are only possible as a function of the pH of the culture medium and could not be conducted except in line with the needs of plants which could be either calcifugous or calcium dependent. In the same way nitrogen needs are not the same for legumes as they are for grasses, etc. The chemist did not see certain aspects of plant biology. This was also the case 80 years later for P. Baranger, who was head of the laboratory of organic chemistry at the Ecole Polytechnique de Paris and did not see certain aspects of physical chemistry such as the importance of photosynthesis and its relation to the incident light spectrum and the materials through which the light passed. He was a doctor of sciences and had done a thesis in chemistry.
FIGURE 1 ~ Photocopy of Nobel Prize nomination
FIGURE 2 ~ Photocopy of Nobel Prize nomination, cont'd.
Chapter 3Several Examples Of Experiments Subsequent To 1974
I will limit this chapter to just those experiments done on oats, referring to my book of 1975 for various experiments on humans, animals (such as mice, lobsters, etc), or with microorganisms, I will briefly comment, in passing, on various results published here and there concerning carbon, silicon, phosphorus, manganese, copper, etc., as they relate to plants. I will not attempt to present in detail various complementary experiments on oats or other plants to study variations of elements such as sodium, magnesium, silicon, copper, etc.
However, I would like to call attention to the fact that studies on the variation of copper in cultures of oats (a plant which is, in the overall, rich in copper) have showed that, with respect to the seed, the plant shows a reduction in the neighborhood of 19% (mean figure from the analysis of five batches). But I judge that we did not have a sufficient number of batches studied to generate a meaningful hypothesis concerning what it is that increases when Cu diminishes (possibly zinc?). My researches on the Mn-Fe link are also not extensive enough although the results tend to converge; however, we must look more closely at reactions in this domain.
I will present in some detail an array of experiments bearing at one and the same time on potassium and calcium in oats in order to show the principal precautions one must take when one comes with naïve eyes into a research area that others have seen from a very different angle. Then I will give in another chapter a few results of research accomplished by J.E. Zundel, research carried out subsequently to those cited in my book of 1975, which he expanded considerably later on and which he carried out limiting himself almost exclusively to variation of calcium in hydroponic culture of oats, in order to show the scope of detailed parameters one must lose sight of in this sort of research. As he always kept me current with his results, I will set forth the core of his research, more especially the memoir that he circulated in the 1979 second semester as a photocopy --- then in an Italian university review in 1980 --- under his signature and consisting of an excellent condensation of 13 years of practically uninterrupted research. Thanks to this huge amount of work focused on a precise area I believe that the study of oat culture has been pushed to a point that we may be certain there is an augmentation of Ca in the order of 100% (varying from 50% to 150% as a function of oat variety, season, etc.), the increase occurring in a plant which has germinated several weeks being compared to a grain similar to the one from which it came without there being any chance that this calcium came from the air, the water, the materials used in the culture equipment, or as an artifact of analysis. Unfortunately, due to human vanity, I understand that it is very difficult to obtain exact replication of an experiment. There are many who consider themselves superior to the rest of the world (or more clever), and they criticize what has been accomplished, desire to generate an experiment modifying the results in line with their own procedures or materials in order to get their own name in the ring. Thats human. But also quite frequently that comes from a misunderstanding of the main problem resulting from distorted professional judgment.
But these inevitable human eccentricities did not curtail widespread dissemination of what some call a "mutation" and what I call a transmutation, a label adopted universally for this physical phenomenon of the mutation of nucleons in atomic nuclei. Thus it is that Prof. Genevois, in the introduction to his Biochemical Mutations of Plants, was able to write a few years ago, "It now seems, from collecting the facts on all cultivated plants of any importance, that the condition of mutation is a general fact. Mutations are in the process of changing domesticated vegetation. Perfecting analytic techniques... that consistency of the composition of plant species was an illusion". One could not say it more clearly, but it is so pleasant to cradle oneself with illusions, right up to ones death!
Now lets demonstrate scientifically that consistency of structure is truly an illusion to be dispensed with!
(a) Research on Variations of Calcium and Potassium in Culture of Oats ~
I elected to continue investigating oats because that is the most calcifugous cereal I have every encountered in my studies. It grows well in acid soils (granites, for example). Accordingly, one can grow it in a laboratory with extra-pure water which is almost always acid (therefore a proton donor according to the modern definition of the acid state --- and we must not forget that which represent the pH of a pH meter). Accordingly one must avoid neutralizing or "stoppering" it. I used extra-pure water resulting from combustion of hydrogen and oxygen, these two gases being the product of electrolysis. They were, then rigorously pure, which is not always the case when, for example, hydrogen comes from a reaction of sulfuric acid on zinc. The zinc is often too impure except when it has been produced by electrolysis. Commercial sulfuric acid is often produced by calcinations of a very complex mixture of pyrites which requires that it be rejected for experimental use unless it is a pure acid coming from refined sulfur. It was not always possible for me to have a sufficient amount of such water; then I used bidistilled water because in water which has been distilled only once a "head" [azeotrope] is formed at the outset of distillation, a condition in which there are very diverse volatile products which condense before anything else --- organic products which are often toxic. My colleague in the Paris Council of Hygiene, P. Levine, of the Pasteur Institute, advised us that he was not able to conduct certain experiments in microbiology or in human biology using the water of Paris with just one distillation. He found it necessary to throw out the "alembic heads" and redistill the remaining condensed portion.
Softened water, purified on ion exchange resins, also called permuted water, still contain too many minerals for our research work. Permuted as well as possible, they must still be distilled by a single passage. Analyses of every batch of water were made by atomic absorption spectrophotometry on samples which varied from one to three liters per carboy and were reduced to 50 ml by evaporation to ensure they did not contain a measurable amount of Ca++ and K+ (since the experiments were concerned with these cations) which could have influenced the results obtained by more than one percent. Such experiments were also carried out on all those materials which might come in contact with the cultured plants (purex, plexiglass, altuglass, polyethylene, etc.). It was also necessary to consider analyses at different stages of the operation to be on the lookout to avoid having parasites get into the circuit.
It seemed to me advisable to cite a few examples of such experiments, according to publications that I distributed in photocopies, for the most part, extracting, however, all the material which was simply repetitious.
(b) Research on K and Ca in Oats ~
I abandoned research on calcitropic plants, like ray-grass, due to a lack of time and personnel, and in the end I limited myself to studies of oats, a calcifugous type of plant. In these efforts I made use of the publications of Zundel and some others. Thus, I could take into consideration a very large number of results showing a statistical convergence which could not be denied by any conscientious person. And very shortly thereafter we find balance sheets on Mg and Si which, under certain circumstances (in animals for instance), both may be able to transform into calcium. Already in 1799 Vauquelin had demonstrated that if silicate diminishes and lime increases when one tallies up the content with respect to a chicken as a function of ingested oats, the diminution of Si does not correspond quantitatively to the augmentation of Ca. Some studies --- perhaps too few --- have shown that in the germination of oats Mg and Si did not play a significant role in the production of Ca. By contrast there were a good many experiments confirming that there was a diminution of K commensurate with the increase of Ca.
Zundel, likewise, only carried out a few studies on Mg and SI in his oat cultures because his first investigations showed him that the variation of these two elements was too slight and insignificant with respect to the increase of Ca. And he also set aside investigation of K for two reasons:
He did not himself analyze K and did not wish to simply play the role of confirmatory of that which had been done by others;
It seemed to him that the various analyses which he has had done by flame spectrometry, by neutron activation, and by x-rays, for example, varied too much as a function of the methods, the operators, the laboratories, and even the operators in one laboratory using the same method. Repetition was not assured with respect to the numerical values of K that were given to him and he had no way, himself, to determine the results which he could trust because he placed his confidence in gravimetric chemical analysis, weighing milligrams of cations. In this way he could measure to one-tenth of a milligram the creation of Ca, but with respect to what? He does not pass judgment on that.
I will not give here results obtained by chemical methods, gravimetric or colorimetric. Rapid analyses for crude verification have been made by various methods of complexometry (by EDTA for example). They are often more delicate to accomplish than one might think because the complexant must be chosen as a function of the affinity between Mg and Ca. I indicated in my book of 1975, with respect to research on the lobster, why it was necessary to reject certain methods because the affinity of Mg/Ca varied a great deal as a function of the metabolism of the animal. After moult of the lobster, the method did not work in the majority of experiments which made it very difficult to obtain results which could be properly compared.
[ PARAGRAPH MISSING ]
I know --- and I have cited some results --- that convergent values for Ca have sometimes have obtained by flame spectrometry. But I cannot have confidence in them a priori; they must be cross-checked, because some equipment uses a flame of a temperature insufficient to ionize enough Ca atoms and obtain valid and reproducible results. It is also preferable to have the same operator in order to better control the regulation of the flame (the delivery and pressure of the gas must be verified regularly).
(2) Simultaneous Investigation of Mg, K and Ca ~
To avoid a long drawn out description in the following paragraphs, I will present only mean data for each whole experiment and will not give mean data for each experimental segment. Obviously, in this experiment it serves no purpose to take account of variation in Mg since that result has been obtained from ten other experiments conducted to obtain balance sheets of Mg when placed in a culture of oats of different varieties. Thats one reason which led me to terminate research on variations of Mg in 1976.
Comments on Analyses of K, Ca and Mg in Seeds and Plants of Oats in the Paper-Mill Analytic Laboratory of the Grenoble University Complex ~
(a) Experimental Protocol ~
Oat seeds of the Flamingskrone variety (a blonde hybrid) with a 27,5 mg mean weight per seed were germinated in special vats covered with an indented plate with two seeds per indentation. The vats received double distilled water with a pH = 5.6.
A preliminary control was accomplished by destroying the water and material of a vat to accomplish spectrophotometry of atomic absorption to verify that there was no measurable amount of calcium therein. The culture was accomplished without added fertilization.
Germination took place in an enclosed space of about 70 x 40 x 30 cm of transparent plastic material out of contact with seeds and water. The enclosure was swept with air sent by an electric pump at a rate of about one liter/minute. This air passed through an air filter provided with one meter of hydrophilic cotton folded and compressed. Then it was muddled through four one-liter glass bottles, each filled with 750 ml of double distilled water, arranged in a series. The first was supplied with 30 ml of HCl to precipitate any trace of calcium dust which might have passed the filter. Then the air passed successively through two bottles with additives of NaHCO3 in order to neutralize any trace of acid drawn by the air from the preceding bottle. The fourth bottle contained only pure water.
Thus, neither by air, nor by water, nor by material in contact with the germinating seeds was there any possibility of an introduction of Ca into the enclosure where there was an overpressure of approximately 3 mm water which made it impossible for any entry of ambient air contaminated by Ca.
At the end of several weeks the plants developed from these seeds were gathered, dried, incinerated at 950 C, dissolved in hydrochloric acid and aliquot parts were analyzed by a number of methods in order to cross check results.
At the same time there were analyzed some control segments of non-germinated seeds as nearly identical as possible to those which were germinated. All seeds utilized were calibrated and came from selected seedings furnished by the INRA and their germination rate was better than 95%. Nevertheless, each seed was hand-picked, the same for the control segments as for those which were germinated, in order to eliminate any abnormality of dimension (either too large or too small) or presenting a visible defect of form or of color, to have experimental segments as homogenous and as similar as possible.
The essential purpose of the experiment, within the framework of verifying my studies, was to compare the quantity of Ca between seeds and plants in order to establish a balance sheet showing that germination of the oat --- a calcifugous plant --- in water with an acid pH will actually alter the quantity of Ca a few weeks of growth in a calcium free environment.
[MISSING TEXT]
...It was requested of the Paper-Mill Laboratory located in the University of Grenoble complex (we note that the University of Grenoble supports the Superior School of Paper Manufacturing which trains the production engineers in this profession) to measure not only Ca and Mg by spectrophotometric atomic absorption but also to measure K by flame spectrophotometry. This was to cross check the results of previous experiments made by Kervran and by Zundel, independently, showing that the increase of Ca discovered in such experiments on oats would only have come from a reduction of K which was quantitatively almost the same.
The measure of Mg was requested of Grenoble because previous chemical analyses of Zundel and physical analyses made by myself had always shown that in the growth of oats, a calcifugous plant, the Mg was practically invariant. One more confirmation of this came to light. I cite results from this Paper-Mill factory done for Zundel who sent them tome for my information. The analyses were accomplished in 1970-1971 but I did not publish my comments except by individual letters in September 1976, with a photocopy of the schedules of the analyses. The following is a combined study.
(b) Tables of Results of Analyses ~
We will only consider experimental segments with identical numbers of seeds and plants, in the two studies, one on Ca and Mg, done first, the other on K, done later, but on aliquot sections coming from the same experiment.
N.B. --- Question marks after a value indicate that it is possible (in my opinion) that there was a slight measurement error with respect to this sample.
(c) Recapitulation ~
The possibility of slight errors from one sample to another is offset by the law of large numbers, the analyses being carried out on hundreds of seeds and plants. The following table recapitulates the above material rounded off to three decimal places. Values are stated in mg
Remark: The variation of Mg in absolute value is very slight ()0.0069 rounded to 0.007). The range of values per plant, for the four analyses, runs from +0.0059 to - 0.0072 whence, around the mean there is a dispersion of 0.0065 (also rounded to 0.007). That is to say, as a function of experimental error, the variation is equal to the dispersion. Fomr this fact we conclude that there is no significant variation of Mg, which had already been confirmed by a number of previously conducted experiments on oats.
(d) Comparison of Variations in K and Ca ~
In absolute value, K diminishes by 0.033 mg per unit while in absolute value, Ca increased by 0.032 mg per unit (between one seed and one plant derived from a similar seed).
Thus there is a definite convergence between these two values, which allows us to conclude that the augmentation of Ca comes from reduction in K, from which we have the following statement:
K 39/19 + H 1/1 >> Ca 40/20 + ~ 0.01 u.m.a.
In different experiments this compensatory augmentation of Ca and reduction of K during germination of oats in an acid culture has been observed to approximately ± 4%, reflecting inevitable differences due to slight biological variations and experimental errors. Again, we can present these values as follows:
in a seed: K + Ca = 0.140 mg
in a plant: K + Ca = 0.139 mgIndicating that the total amount of K + Ca does not change. or even:
in the seed: K/Ca = 0.113 / 0.027 = 4.2 approximately;
in the plant: K/Ca = 0.080 / 0.059 = 1.4 in excess,which again reflects the reduction of K with respect to Ca. As K + Ca does not change, the increase of Ca can have no other origin than the diminution of K, which makes it useless to look for some other origin of Ca. However, as we find certain metabolic systems (especially animal systems) in which Ca could come from Mg --- by the reaction Mg12 + O8 >> Ca20 --- due to complimentary adjustment, research on variation of Mg was made one more time, in the experiment commented on herewith, and it was discovered to be non-existent. Likewise, other previous experiments had shown that there was no significant variation of Si during the germination of oats. Bear in mind that a calcitropic plant does not manufacture Ca, which must be brought to it by the culture medium.
The values of K and Ca, in both seeds and plants, as set forth above, are accepted without reservation because they confirm the mean measures supplied by other investigators using a variety of different analytic techniques.
It is noted that in this experiment the relative increase of Ca is greater than 118%. The graph in Figure 3 clearly shows the reciprocal variations of K and Ca.
(e) Review ~
For purposes of comparison let us consider a previous analysis by neutron activation made by a Swiss government center for nuclear research on seeds of a closely related variety, Peniarth (mean seed weight 32.2 mg). Analyses were made on five samples of six seeds and five samples of six pants each.
Figure 3 ~ Inverse variations of K and of Ca in germination of an oat seed (Mg does not vary significantly).
Mean figures per seed, from 3 analyses and a total of 840 seeds.
Mean figures per plant, from 4 analyses and a total of 403 plants.
Analyses of K by flame spectrometry
Analyses of Ca and Mg by atomic spectrophotometry absorption
Analyses made at the Laboratory of Paper Products Analysis (University of Grenoble).
DK = 0.113 - 0.80 = -0.033 mg
DCa = 0.059 - 0.027 = +0.032 mgTable 3: Batches of 6 Seeds and of 6 Plants (Values in mg)
So again we have an augmentation of Ca in a plant, compared with the Ca in a seed similar to the plants seed of origin in the amount of 0.0560 - 0.0249 = 0.0311 which gives an increase of 124%. We are not dealing here with the same plant variety as in the preceding experiment, but we see that the order of magnitude of variation remains very close to the same. Actually, the values obtained in Switzerland were corrected some time after issuance of the first analyses because it was noted that, because of some germination failure the values reported applied not to a total of 30 plants but rather to an average of 28.32 plants, thus leading to a slight increase in the amount of Ca calculated for the plants. Another calculation was added, by comparing the content of Ca, not just by the unit (seed or plat) but as a function of weights and the values were given in ppm, which allows us to compensate the inequalities of weights of the samples to obtain a more homogenous result. This led to confirmation of a 138% increase of Ca.
We can briefly add other results showing the convergence of values obtained by different laboratories, always with oats:
1. Nuprime variety, Kervran culture, 42 days; analysis by Dr Bieselaar, former lab director of Fraud Service ---- Atomic Adsorption Spectrometry; average grain weight 24.65 mg (dozens of experiments with this variety have given average weights from 22 to 25 mg, according to the source of the lots).
2. Flaningskrone variety; average seed weight: 27.5 mg (has varied from 25 to 30 mg according to the source. Ca/seed: 0.028 ~ Ca/plant: 0.056 ~ Ca change: +0.028 = 100%
3. Peniarth variety: using neutron activation, 52 day culture; Ca/seed: 0.057 ~ Ca/plant: 0.052 ~ Ca change: +0.0263 = 103%
4. Same variety, dosed by atomic adsorption spectrometry, after 48 days of culture: Ca/seed: 0.0260 ~ Ca/plant: 0.064 ~ Ca change: +0.038 = 146%
5. Peniarth variety; using neutron activation, Ca/seed: 0.0249 ~ Ca/plant: 0.0572 ~ Ca change: +0.0323 = 130%
All these examples definitely show similar results and an average increase in Ca greater than 100%, which is absolutely beyond all possible experimental error and confirms the "creation" of Ca, pointing to a transmutation. We have seen that it begins with K. Dozens of experiments, made up of hundreds of analyses, performed by different laboratories using varied methods, have been conducted on tens of thousands of oat seeds and plants. The phenomenon of transmutation is therefore well established beyond doubt. The increase of Ca tends to become asymptotic after 5 days too 6 weeks of culture, which implies an exhaustion of synthetic hormones during germination.
A transmutation by a lining organism does not follow the classical rules of high energy interaction. Instead it can be classified in the category of low energy interactions. The loss of mass is not explained by a release of heat and is not represented by any radioactivity --- alpha, beta, or gamma. In my 1975 book, Costa de Beauregard, Research Director at CNRS, showed after the confirmation of "neutral currents" in 1974 that the loss of mass is explained by introducing the action of neutrinos. Therefore, the loss of mass does respect the classical theory of physics. I have added to it the action of the neutral intermediate virtual boson vector z, and the previously stated reaction could be written as:
C + K + H +Zo / + enzyme >> Ca + v1
with v prime not equal to v pertaining to energy level. The introduction of the H+ proton would be explained by the tunnel effect by application of quantum mechanics.
In the above reaction v is different from v from an energy point of view. The entry of the H+ proton would be performed by the tunnel effect in accordance with quantum mechanics.
The text just quoted was released in French from September to October 1976. It was soon after translated into English and released in the USA by Dr E. Stanton S. Maxey, surgeon and owner of a surgical-medical center in Stuart, FL, who has done much for the release of my research.
The above stated results seemed to me definite proof of an increase in Ca by an equal reduction in weight of K. I saw, therefore, more or less, remarkable support to establish the reaction proposed for approximately 15 years, given by the formula K + H >> Ca, resulting from a great number of observations which all tended to converge on the same conclusion. I attributed the slight variations of the numerical results to cultural protocol; to slight differences in the strength of germination of the seeds; or to non-eligible, hard-to-measure cosmic phenomena that have been confirmed experimentally by numerous scientists, such as the influence of seasonal changes, etc. Subsequent experiments came to confirm such results.
FIGURE 5 ~ Formation of calcium from potassium by the combined action of an enzyme and a neutrino.
The positive charges of the active site of the enzyme repel the H+ proton. The result of this electrostatic field is represented by H. H does not appear as a label in this figure. [Ed. --- I think he means the result of this electrostatic field is felt by H, as shown in the figure].
The enzyme would concentrate the neutrinos v, increasing the chances of impact with the material.
A neutrino v, adding its effect to the enzymes positive charges, repels the H+ proton towards the K nucleus with sufficient energy for the proton to penetrate the K by the tunnel effect. The K atom recoils a little from the shock and becomes Ca (K19 + H1 >> Ca20).
The incident neutrino v, which has accompanied H, does not penetrate very far into K. It has given up some energy to H and is refracted in K, leaving with a different energy v prime not equal to v by carrying off the excess energy resulting from the loss of mass between Ca and K + H.
This re-emitted neutrino v prime will be lost in space without reacting with the material.
But I have never been able to understand why certain experiments performed elsewhere seemed to indicate that there was not always a one-to-one correspondence in the increase of Ca to the decrease in K, the loss of K being insufficient to account for the increase of Ca. There was not sufficient reduction of Mg or Si to compensate for the increase of Ca. Nor did I see any evidence that the analytical methods could be blamed. Was I to infer from this that it is probable that we have not yet completely mastered the cultural protocols that allow us to obtain reproducible results of the K content?
But it was no longer possible for me, for diverse reasons, such as age, health and financial means, to start over again with a systematic study of the K life cycle, which would take years. I therefore decided to continue only with the research that always pointed to an increase in Ca, since Zundel himself was not analyzing K, but was instead specializing his research on the increase of Ca in oats. There was a restricted but sufficient domain here to establish the increase in calcium, and therefore the creation of matter in the germination of oats. That is to say, there is a biological process of transmutation of matter, and that there are other phenomena besides chemistry in living things where there is no evidence of high energy transmutation. But I thought for the time being that we needed to be conservative about the role of K in the formation of Ca. Researching the element, or elements originating from Ca is quite another problem that young scientists will have to solve in the future.
FIGURE 6 [Not included here] ~ J.E. Zundels Research Station in Grasse, France (1976) --- Environmental culture tanks. Pumps and purification flasks that control the air sent into the tanks are in the back room.
First, I will present a study by Zundel that he had published in September 1979. He only measured the calcium content, as you can see the results in his summarized table, which is reproduced also. Then I will go on to some commentaries on diverse experiments, with the logical conclusions that can be drawn from them. The following text has been reproduced in French and Italian in Rivista di Biologia (Fall 1980), Univ. of Perouse, directed by Prof Sermont.
(3) Research by J.E. Zundel ~
I introduced this researcher in my book of 1975. J.E. Zundel graduated from the Zurich Polytechnicum as a chemical engineer. He then trained in the USA. He managed a paper mill, keeping for himself the supervision of the chemical analysis laboratory. This allowed him to maintain his proficiency in the field of chemical analysis for the rest of his life. He had a completely open mind and he was a deft experimenter. My first works were a revelation to him. He wrote to me as early as 1963. He understood immediately that my works pointed to a whole new aspect of biology which was of great interest for pulp and paper research or on all organic material obtained from vegetable fibers. Some alterations in paper could not be explained by chemistry alone. Micro-organisms would implant themselves in the paper and cause modifications which could not be understood. In 1963 Zundel began some preliminary verifications of my ideas after broaching the subject with me in letters. His work was intermittent, because his professional activities did not leave him the tie required for continuous research. However, this preliminary survey convinced him that my works contained a reality, so far ignored by all chemistry treatises and specialized periodicals. As soon as he retired, he moved to a pollution-free location in the country, half way between Gasse and Canes. At his own expense he installed a hydroponic experimental station housed in a Vitrex greenhouse. Vitrex is made of fine wire mesh dipped into pure Cellophane, a solution of pure transparent celluslose. Vitrex is more permeable to the solar spectrum than glass. It is well known that glass filters some ultraviolet wavelengths. Vitrex is selective to several infrared wavelengths which are conductive to the greenhouse effect. This Vitrex greenhouse also protected the plants against unavoidable small dust particles. Some dust was still brought in and out of the greenhouse on shoes and clothing despite these precautions. Zundel even worried about dust carried by the wind from a quarry located 10 km away. This was indeed improbable, but it had to be considered to counter any subjective objections raised by the eternal systemic objectors. To present accurate digital data, he interspaced his planters with control planters filled with pure water. In this way he could accurately measure eventual dust fallout. In addition, he equipped a chemical analysis laboratory for the measurement of Ca and some other elements such as Mg and Si. He personally did the various analyses, as he distrusted lab technicians (I appreciate these highly qualified technicians. They are very useful collaborators. Nevertheless, one should keep in mind the old saying: "Better deal with God than with his saints", especially in a new field, which has not been taught to them). This hobby kept Zundel busy during his retirement.
It is important to note that Zundel never measured K himself. This element is difficult to measure by purely chemical methods. At Ecole Polytechnique in Paris, Prof; Baranger. Director of the Organic Chemistry Laboratory, also abandoned the various chemical methods prescribed in classic analytical treatises after he had performed questionable chemical analyses of the element K. Subsequently, he used a physical method, fashionable at the time, in order to avoid, as he told me, malicious comments by his "dear colleagues", always ready to find faults in procedures set up by others. Only their method is reliable. I will not describe the multiple causes of uncertainty in chemical methods. They are usually related to a sequence of the solutions ending up with a compound which is insoluble in the next phase. Too many chemists forget, in their quest for an element inside a seed, that this element is related to different compounds in the plant grown from this seed. They reason solely as chemists and they forget --- or ignore --- biological phases. For a long time now, analyses of K have been performed essentially by flame spectrometry techniques (emission spectrometry) which have been refined and automated. These techniques are easy to set up with a butane or propane flame. However, preparatory steps remain delicate for flame spectrometry as for other physical methods.
When Zundel wanted values for K, he usually would send ash samples to a well equipped Industrie du Papier laboratory linked to the Ecole Superieure du Papier of the University of Grenoble. In these cases, he only mentions the results without lending them his support. He only claims as his own the data which he obtained himself and checked by a different method. This is the case for calcium which he measures following a gravimetric method. The samples are weighed on a high precision Sartorius scale sensitive to the 1/10 mg. This is sufficient considering the quantities involved. Baranger weighed his sample on a Mettler scale sensitive to 1/100 mg. Such precision is meaningless, because it is subject to gravimetric variations such as the distance to the operator. Most recent models were calibrated on very heavy bases to minimize the influence of distance to the operator. Still the operator should not get too close to the scale and he should use proper remote controls. Protections are necessary to shield the scale from the operators body heat and breath.
Zundel performed tens of thousands of experiments, including hundreds of analyses on tens of thousands of seeds to establish a procedure guaranteeing reproducibility. He wished then to check to results obtained using the method recommended by Prof Charlot of Institut de France in his now classic Traite dAnalyse in order to eliminate any possibility of error. After preparing a sample (an aliquot part) of ashes himself, he had it analyzed with an atomic absorption spectrometer at Laboratorie des Industries de la Papeterie in Grenoble. It was a Perkin-Elmer instrument. I used a Beckman. He had the concentrations in Ca and also in Mg, obtained by Charlots method, tested with this instrument. We will not discuss these concentrations here, as the variations in Mg proved to be small, if at all significant. The results obtained by Zundel on oats regarding Mg confirmed mine. I stated this point in my book of 1975. This is not the case for the amount of the variations of Ca in oats, a calcifuge plant. In calcicole plants the metabolism of Mg is totally different. I will not elaborate on this subject and I will limit my topic to a wealth of studies. In this field it is not permissible to extrapolate results to other plant species without having a very large number of data taking into account the numerous parameters which should be considered in the growing process.
Zundel used a third method, neutronic activation, in order to check his data. He entrusted these analyses to the Institut Federal de Recherches sur les Reacteurs Atomiques located near Zurich. As Zundel could cross-check the data by three totally different methods, he was able to select the experiments which gave three sets of compatible figures. Any large discrepancy in one of the figures would lead one to suspect an error in the analysis by the operator. In this way he happened to detect a dilution error in an experiment with the atomic absorption spectrometer. Ashes were dissolved in hydrochloric acid and then diluted in twice distilled water prior to injection into the instrument. The analyzer was programmed to average data, measured automatically on ten samples contained in small test tubes. In this analysis there were obvious variations in the results as compared to those obtained in a large number of previous analyses. The operator made the mistake by one order of magnitude. Sometimes the error is due to an oversight. For example, something sticks in the bottom of the crucible after firing. A calibration error is another possible cause, so is a zero displacement. These are some of the reasons why one cannot rely on a single experiment.
I will mention again that I performed germination experiments on two layers of ash-free filter paper in Petri dishes. I measured the total Ca content of plants and paper. In another experiment, seedlings were cultivated in the cells of a seedling tray which had holes at their bottoms. Operators had neglected to measure the Ca which migrated through the roots to the underlying water. It was only an error of secondary importance. I ascertained that this Ca contained in the water (null at the start) represented approximately 10% of the Ca increase in the plant as compared to the Ca content of the seed from which it originated. This increase was often greater than 100%. However, to be rigorously scientific, it was necessary to take into account the Ca content of the water. I informed Zundel of the necessity to perform such measurements, which were omitted in the first experiments.
In my book of 1975 I only mentioned Zundels studies prior to1972, at which time he presented a paper to the Academie dAgriculture de France. Zundel did not publish from that time until 1979. He pursued his study continuously during the whole period. I did not want to publish the results he was sending me all along, in order that he would be the first to do so. Also I only wanted to comment on data that he had published himself so as to have a public database, which would not be the subject of objections. I will reproduce below a text printed in 1980 in the Rivista de Biologica (quarterly issue, 3rd quarter 1980), published by the University of Perugia. This is the original text followed by my short commentary which is my sole responsibility.
FIGURE 7 ~ Hydroponic Culture of Grains
"The planter, presently made of polyethylene, includes a tray in which twice distilled water is maintained at a fairly constant level. The pH is 6.5. The water level is topped off every 3 or 4 days to compensate for plant transpiration. This is done by means of a fixed tube linked to the outside by a siphon. The external extremity is covered with a test tube as a hood for dust protection. The test tube is only removed to introduce the pipette for water additions. A multi-cell panel, also in polyethylene, is placed on top of the tray. The 75 cells receive two seeds each. The roots reach for water into the tray below, wile the seeds are maintained outside the water (aerobiea). A maximum of 150 seedlings can be grown in each planter. Two chambers, as the one reproduced below, are fed in parallel from a same air supply.
"The planter is placed in a closed chamber made of thick, rigid Plexiglass 70 x 40 x 30 cm. The box is closed with a removable Plexiglass panel after introduction of the planter. This panel is fixed by steel clips to one side of the box. It is screwed to the box once everything is in place. A gasket insures air-tightness between the panel and the box.
"Air is pumped into the chamber at an approximate rate of 1 liter/minute by a device which is not represented on the figure. The air is pushed through an air filter by an electric positive-pressure pump. From the filter it passes successively through four one-liter flasks partially filled each with 750 ml of twice distilled water. 30 ml of HCl are added to the water of the first flask to precipitate any dust (Ca, Mg, etc.) which might have escaped through the filter. A check showed that the filter was effective. The air is scrubbed in the next two flasks filled with solutions of NaHCO3 in order to neutralize any acid carry-over. A final scrubbing is made in the fourth flasks which contain only pure water. From there are filtered and scrubbed air penetrates into the cultivation chamber. Air exits the chamber through a double siphon, used also as a water gauge. A positive pressure of 3 mm W.G. was measured inside the chamber. This way, no external air intake may happen accidentally, even with a defective gasket. No Ca, for example, can be brought from the outside".
(4) Study of the Variation of Calcium in Oat Seedlings During Germination in Twice-Distilled Water, by J.E. Zundel ~
Abstract: In the course of 60 experiments the Ca content of oat seedlings increases by 50-250% during germination in twice distilled water.
Introduction:
(1) This research was carried out over the last 13 years, following the publication of works by Monsieur C. L. Kervran. These works suggested a possibility of mutation in chemical elements by biological means, in contradiction to the law if immutability of matter posited by Lavoisier, being understood that the law remained fully valid from the chemical point of view. Kervran was kind enough to follow my work with interest. His considerable knowledge was for me a source of frequent and effective advice.
(2) It was by mistake that I selected oat for my study. Initially I was looking for silica in a study which was not pursued. Even though the oat was a calcifuge plant, it showed a strange increase in Ca which was to become the subject of this study.
(3) It was obvious that this study required the elimination of all Ca additions of external origin during the experiments. This, I tried to accomplish over the years.
(4) During the presentation of a paper of a paper to the Academie dAgriculture de France, I was strongly criticized for deleting experimental details. I will present them here as accurately as possible.
Growing Procedure:
(1) At the beginning of my work, I was content to use common, fodder types of oats. Later the Centre des Semences of INRA kindly supplied me with selected seeds: Nuprime, Flamingskrone and Peniarth, species I finally adopted definitely for its high germinating power. The Warburg and triphenyl-tetrazolium-bromide tests showed a germinating power of close to100%.
(2) After trying several methods (beakers, Petri dishes, flat and hollow plates) I selected Mutipot seedling trays, manufactured by Ossenberg.
Each device included a tray ( 30 x 50 x 3 cm) with a panel of the same dimensions with 73 cups 40 mm deep which were perforated on their bottoms. The assembly was made of PVC. The analysis of the trays showed no Ca.
(3) The seeds were sized by sight with a ± 3% accuracy in weight.
(4) For pre-germination, I cover the bottom of a Multipot planter with two layers of ash-free Whatman filter paper and I saturate the paper with twice distilled water. I spread over it 300 selected seeds and I place the assembly in a phytotron. Temperature is controlled at 28° C during the 16 daylight hours and to 15° C during the 8 night hours.
During the day, the phytotron is lighted from the outside by a 400W Power Star Osram lamp. After 5-10 day pre-germination, the seedlings are 30-70 mm high. The duration of the pre-germinating phase, as well as the size of the seedlings, seem to show that besides controllable factors, there are other effects which escape me. I then assemble the two parts of another seedling tray and I fill it with twice distilled water to a level 4 mm above the bottom of the cups. I pull the seedlings carefully not to hurt the radicles, and I transplant them two by two in the tray. The complete tray holds 146 seedlings. I place the tray in the phytotron.
(5) The seedlings are harvested 28 days after the beginning of pre-germination. I selected this time span because it corresponded to the average duration of the plants, hence probably to the reserves in the seed. I pull the plants carefully out of the planter (roots may extend as much as 20 cm between the holes at the bottom of the cells and the underlying tray. I spread the plants on Whatman filter paper and I dry the whole thing at 85 C for 72 hours in an electric autoclave. I then grind the plants in a Moulinex coffee grinder and I weigh them.
The water used for the growing process (1,500 ml in average) is recovered and concentrated to 50 ml for spectroscopic analysis.
Elimination of Extraneous Ca:
(1) The twice distilled water was supplied by Laboratoires Aguettant in Lyon. 3000 ml of this water, reduced to 50 ml, do not show any Ca on the Perkin-Elmer spectrometer.
(2) The glassware is made of pure silica.
(3) The crucibles are made of platinum, with the exception of the ones used to incinerate seeds and plants. For this work I used crucibles of pure silica.
(4) Reagents were supplied by Prolabo and were of analytical quality.
(5) From the beginning of my work, I was afraid of the Ca brought in by ambient air.
Therefore, I placed a tray with the same dimensions, filled with HCl, N/10, next to the culture for the duration of the experiment. I recovered amounts of Ca corresponding to 2-3% of the amounts measured during germination. As an additional precaution, I enclosed the seedling trays in polyethylene cases and force ventilated with clean air.
Later I had airtight cases made of Altuglass which also had to be ventilated. Finally I acquired a fully airtight phytotron (140 x 60 x 50 cm). Its case had a lid made of special plexiglass, more permeable than glass to wavelengths below 400 mm. Lighting was provided by a 400W Power Star Osram lamp located 45 cm above the plexiglass panel. This lamp emits a fairly uniform spectrum extending into the ultraviolet. The light intensity at plant level was approximately 5,000 lux. I some experiments I added a Mazda Ultraviolet 20W lamp.
A small compressor provided air ventilation (2 liters/minute) for the phytotron. This air was first filtered in compressed pharmaceutical cotton. Air purification was continued by scrubbing into two flasks (2 liters) filled with HCl, N/10, one flask of concentrated NaHCO3 and one flask of twice distilled water. Following some objections, I replaced the cotton filter with a Whatman Gamma 12 filter of 0.3 micron porosity. I did not find any significant difference in Ca fallouts using these various protective methods.
(d) Analyses:
(1) Samples were weighed on a 0.1 mg Sartorius scale.
(2) Seed and plant incineration was made in closed silica crucibles on low Bunsen flame.
After distillation, I place the crucible in a slanted electric furnace heated to 800 C. Some objections were raised about this temperature. However, my own experiments and other experiments performed at Laboratoires Techniques du Centre Technique du Papier in Grenoble, showed no appreciable differences for temperatures of 550 C, 600 C, 800 C, 1050 C.
(3) Silica was recovered from the solution following the Treadwell method. The precipitate incinerated in a platinum crucible was weighed and diluted in 5 ml of concentrated HF. After vaporization the SiO2 amount was obtained by weight difference.
(4) The determination of Ca (and its separation from Mg) was performed by Prof Charlots method, as described in his Traite de Chimie Analytique Minerale.
(5) The determination of K was performed by flame spectrometry on a Perkin-Elmer instrument at Laboratoires du Centre du Papier in Grenoble.
(6) All data were expressed in milligrams per unit of air-dried seeds or plants.
(7) Some result data was cross-checked by atomic absorption spectrometry at Laboratoires du Centre Technique du Papier in Grenoble. Generally the values obtained with this method were close to my results, although I noted discrepancies as great as 30%. Nevertheless, percentages of increase in Ca between seeds and plants were similar to those I had obtained.
Additional checks were performed by neutronic activation at Centre de Recherches Nucleaires in Zurich. Lesser discrepancies were noted, but percentages of increase in Ca were equal to mine.
Results:
(1) All data the least bit suspect of Ca contamination from the air was deleted from the table. Consequently, I only mentioned the data obtained from experiments in the phytotron and in the Altuglas, polyethylene and Vitrex cases.
(2) As expected plant weights were greater than seed weights (photosynthesis) but in an irregular fashion. There were two exceptions: item 345, phytotron, in total darkness, and item 353, Altuglass supplied with air without CO2 (consequently there was no photosynthesis in either case).
(3) Ashes increased. However, item 329, phytotron, was an exception. Ashes decreased for no obvious reasons. In seeds, P was included in organic compounds (phytins) which disappeared during incineration. In plants we found P as a non-volatile Ca-phosphate. The other source of weight increase of the ashes, was the formation of Ca. Probably from C?, i.e., 2C + O >> Ca or 2 x 12 = 16 = 40). It should be noted that the increase in ashes is not proportional to the increase in Ca.
(4) Mg remained identically the same. SiO2 varied slightly in either direction.
(5) Ca showed a 52-292% increase. Here was one exception for item 342, phytotron, additional lighting with an UV lamp and addition of CO2 in the air supply, for which the increase in Ca was 556%. This result was checked by spectrography and neutronic activation. I could not duplicate this result under identical conditions (item 345). This test showed only an 88% increase. There was an additional exception for item 264, no Co2 and no UV. There was certainly a mistake, but I could not locate it. I mention these two experiments as I want to be absolutely candid.
(6) Spectrographic analyses of the water from the culture at the end of the growth period showed an average Ca content of 0.015 mg per plant. Normally I should have added this amount to the Ca measured in the plants. I did not do it, because a water analysis was not performed for each batch.
(7) K contents in the plants and in the seeds were different. The average variation was of 10%.
(f) Dead Control:
Tests on dead controls are fairly difficult as there are scores of methods to kill a seed. In every case 24 hours at 100° C is deadly. 24 hours at 88° C seems to be marginal, as shown by successive tests at temperatures progressively approaching 88° C. After germinating the seeds for one month, there was no sign of life. Ca and SiO2 were the same as in the fresh seeds.
I tried to kill the seeds with formaldehyde as an alternate method. Eight days immersion in formaldehyde were required to prevent any germination. There again, Ca and SiO2 were the same as in the living seeds. I pursued the tests with gaseous formaldehyde. After a 7-day treatment with the gas, I began cultivation. After 30 days there was still no germination. The Warburg test was negative. However, the test with triphenyl-tetrazolium-bromide was not 100% negative. Therefore, a trace of life remained. Ca had increased by 24%, silica was unchanged and ashes had increased by 24.4%.
Finally, I tried a fourth and more drastic procedure. I ground the seeds. I wetted the grounds and left them in a calibrating glass for 28 days. The ashes had increased by 25%, SiO2 was unchanged, and Ca had increased by 12%.
This test was repeated under identical conditions. It resulted in a 17% diminution for the ashes, in no change for SiO2, and in a 23.5% increase for Ca.
To me, these tests do not look conclusive.
(g) Reproducibility:
I entertained great hopes for better reproducibility from air-tight lids and from the purification of the air supply after obtaining ill-assorted results over the first years of my study. This hope was only partially realized.
The most scattered results were obtained with the phytotron, though it provided constant temperature and relative humidity, and purified air supply.
Item 326b: 111%
Item 329: 52%
Item 358: 64%
Item 360b: 204%These results are indeed disappointing, but I cannot estimate the effect of the parameters influencing the growing process besides those which I could control.
On the other hand I could correlate the results of the three other groups:
(a) Altuglass:
Item 326a: 257%
Item 351: 271%
Item 343: 260%(b) Vitrex:
Item 254: 115%
Item 260: 106%
Item 274: 170%
Item 271: 97%(c) Cultures with no photosynthesis:
Item 315: 54%
Item 345: 92%
Item 553: 84%Therefore, the results were reproducible under some conditions.
These results appear to show the influence of light. The Altuglass, more permeable to light, gives higher results than the Vitrex under solar exposure. Another corroboration can be seen in tests 360a and 360b performed simultaneously:
(a) additional UV; increase in Ca: 286%
(b) no additional UV; increase in Ca: 204%Conclusion:
(1) There is always an increase in Ca during the germination of oats in twice distilled water.
(2) It is likely that this increase comes from carbon after the pattern: 2C + O >> Ca.
It seems also likely that this process is produced in two phases: the first being supplied by the reserves in the seed, the second by photosynthesis.
(3) Light seems to have an important influence, in particular wavelengths below 400 nm.
(4) Beyond controllable growing conditions one must probably accept other influences, yet unknown to me.
(5) It is regrettable that my means (laboratory, time and age) prevented me from performing a series of experiments instead of a single exploratory experiment. It would have been possible to analyze the data mathematically. Certain discrepancies could have been explained, such as the influence of the location of the planets during germination, a subject being researched by several universities.
Grasse, September 1979
(5) Comments on a Study by Zundel on the Increase of Calcium in Oats During Germination in Twice Distilled Water ~
The point of paramount importance in this study, issued during the summer of 1979, is the fact that each one of over 60 experiments shows an increase in calcium in oats, germinated in an environment devoid of calcium. This increase varies between 52 and 292% according to Zundel, and is always of an order of magnitude sufficient to rule out statistical errors.
This test is a summary. It is very short and can be quickly read by all. It reports data for only 19 experiments, including one on wheat, which I will discard. It is one of a kind and I do not have enough precise information from research to establish valid comparisons, It would be tiresome to review the separate results for all 60 experiments.
It is obvious that this text shows a long experience with chemical analysis on the part of the author. Furthermore, his high scientific integrity makes him mention experiments giving unequal results. This is really the value of this impartial study: it is not the result of an arbitrary data selection. The text opens the way to a wide discussion because it presents the research in all its complexity.
The author does not orient his research to obtain a reproducibility at all costs. He wants essentially to vary the operating conditions. In spite of this, the increase in Ca is always large. But what is the source of these variations in amplitude? One must look more closely at the factors which differentiate the experiments.
It is seen that the increase in calcium is significantly smaller when carbon dioxide is eliminated, inhibiting the photosynthesis metabolism. However, too few experiments without or with an excess of CO2 were performed to warrant definitive conclusions. The addition of CO2 is very difficult to control. Small amounts must be continuously injected. Nevertheless, various other parameters allow one to see the importance of the photosynthesis metabolism.
Lets compare the dry weights for the seeds and for the plants. The photosynthesis cause the synthesis of various carbohydrates (glicids, parotids, lipids) which together with the CO2 derived from the air, produce cellulosis, starch, etc. This results in an increase of dry matter in the plant. If no increase is observed (as is the case for items 346 and 353), it must be inferred that the pant lived only on its reserves in carbohydrates, which were promptly exhausted. Experiment No. 345 (darkness) had to be interrupted after 10 days and experiment No. 353 (no CO2) after 16 days because the plants wilted. Due to the abridged growth, the increase in Ca was less than 100% in both cases.
The photosynthesis metabolism is affected by the light spectrum which penetrates inside the transparent chamber housing the culture. Glass is not the best material. It is known for filtering some wavelengths in the ultraviolet. The production of Ca is increased if a special UV light is placed inside the chamber (experiments 341 and 361a). On the other hand, results for no. 347 remain unexplained. This confirms once more that a single experiment cannot be relied on in all its details. Did anything escape our attention? Zundel acknowledges this point when he writes: "besides controllable factors, there are other effects which escape me".
Anomalies of many different origins can happen. This is the case for wheat, for which the dry content decreases in the plant though the amount of ashes increases. The same happens in experiments No. 346 and 353. Sometimes the differences may be due to an oversight; some material sticks to the bottom of the crucible, for example, and it is only detected during the next experiment.
Some increases in Ca may appear abnormal (experiments No. 264 and 341) which does not necessarily mean that there is an error in the analysis. Regarding No. 341, Zundel comments on this aberrant increase, "as I want to be absolutely candid". He cold not determine the cause of the aberration. The data obtained by Zundelin his own chemical analysis was confirmed by compatible results obtained in following two different methods, atomic absorption spectrometry and neutronic activation. His shows that it was not an error of analysis, which is always possible when checks are not made by alternate means. Was the analyzed sample abnormally rich in Ca? Why? It is not possible to answer this question. Was it due to an accidental contamination during the handling of the sample? This is not proven. However, I think that such data should only be accepted with reservation.
In these two experiments it looks obvious to me that all this calcium could not come only from a transmutation of K (not measured in these two cases to my knowledge). On my side I have data on several hundred analyses showing that in one oat seed there is not enough K to account for such a weight increase of Ca. Could it come from Mg? It looks unlikely, judging from past research. From silicium? I do not think so because of various experimental reasons. Zundel thought of another cause (2 C nuclei rotating around an O nucleus?). I believe that no experimental research was ever done along this speculative hypothesis. This shows, once again, that one experiment or even two do not warrant conclusions, even when the experimenter is highly qualified. These 616% and 556% increases in Ca remain an enigma.
There could be some cosmic influences yet undetected. Russian Prof Dubrov of the Research Institute of the Globe, Moscow, mentioned in one of his books a large variation of Ca in synchronism with a large geomagnetic field variation. The geomagnetic field itself varies with solar proton showers. Such causes are diverse, complex, direct or indirect and above, little studied.
Zundel was intrigued by the 556% increase in Ca, which corresponded to additional CO2 and UV light (Ref.341). Is this why he mentioned having duplicated the conditions (Ref. 347) --- except perhaps the cosmic conditions? He only obtained 88% of increase in Ca. According to him, "there is certainly a mistake" in one or both of the experiments. No conclusion can be drawn; in any case, this example raises a wealth of questions. The importance of the filtering of the light spectrum by various materials should be noted. Altuglas would appear to be more efficient than Plexiglass.
Zundels study presents many enlightening aspects and this is why I take the opportunity to comment on it. A commentary always embodies a personal point of view. It is subjective but necessary, I think. Zundel made sure he presented a totally objective work. It was only right, I think, that I showed the reader the lesson to be derived from it, to facilitate the thinking process and to aid future experimenters attempting to duplicate the experiments. These experiments showed a generalized increase in Ca, slightly larger than 100%, after a few weeks cultivation in extra-pure water of approximate 5.7 ± 0.1 pH (in my research). Zundel does not state the pH of the water he used, but I know that it was approximately equal to this value.
Note: Zundels table shows only the variations of Ca. All these data have been checked by alternate methods and they all point to the same conclusion. Without quoting data in absolute value, Zundel states that average K variations between plants and seeds can reach10%. In fact, he had too few data, too widely scattered, to attribute a significant value to this figure from the mathematical point of view. For K I have the data of many other experiments. The uncertain mature of statistics should be kept in mind; after all, statistics measure only "probabilities". Too often one tends to forget it. Statistical computations varied in time and space. Today Gauss curves are little used. In France the fashionable method is Fishers. This method introduces the very arbitrary "Students-t". For it, Prof Baranger had chosen a value of 1; I chose a value of 5. This means that I discounted all variations smaller than 5%, as Baranger accepted variations greater than 1%. In Russia the Kolmogorov-Smirnovs tests are used: in other countries it may be the Kuiper-Stephens method, etc.
(1) About the Dead Control ~
An objection was that biological transmutation should not be observed in seeds which had been killed in dead controls. In fact, significant, though small, variations were observed very often in the mass of several elements when dead seeds were placed in water just like the seeds to be germinated. Therefore, some transmutation could not be ascribed to biological effects and some people were convinced there were errors in the test procedure.
Zundel also observed the same thing and he mentions the subject in his text. However, one observation directed him to a possible explanation of the phenomenon: it was noted that dead seeds laid in water for several weeks produced a cheesy small. Consequently, ferments, yeasts, or micro-fungi had developed on the dead material. The transmutation observed, leading to an increase in calcium, might be attributed to the action of this microflora. This had to be verified.
For this reason, an antibiotic and fungicidal product, kanamycin, was added in a ratio of approximately 10-3 mg/liter. This concentration was recommended by Montuelle and Ochin in 1967 to prevent the formation of mold on germinating seeds. The presence of mold introduces an uncertainty. Should the result be ascribed to the action of seeds or to the action of the mold? Tests were made on the European type of oats, Peniarth. In the USA, fungicidal tests were performed on the local type Montezuma. The fungicide Ceresa was used. All Ca increase in the dead controls disappeared.
(2) Cosmic Effects ~
Zundel mentions "unknown parameters" to explain quantitative variations. In a figure in my book of 1975, I stated that I started the germination process at the new moon and that I harvested 6 weeks later at the full moon. I did this in order to operate (hopefully) under comparable cosmic conditions for each experiment though I could not establish this point in any significant manner. At least for certain plants, the action of the moon is well known among farmers and gardeners. However, it was not clear for oats. The biological transmutation seemed to depend on other causes which I did not have the time to isolate. The action of the moon on tides is well known, but its effect on oat seemed to be variable.
A book published by Robert Frederick around 1980, LInfluence de la Lune sur les Culture states that oat is an exception, that it might be more sensitive to some cosmic rays. However, it does not appear that the author tried to determine which ones. One must also consider solar action, which shows its effects on high equinoxial tides. The effect of solar proton showers was studied in particular by Prof G. Piccardi of the Institute of Physics and Chemistry of Florence, Italy. In Russia this effect was studied by Prof Dubrov of the Globe Geophysical Institute of Moscow, etc. The research should be pursued for specific vegetal species and it should not be limited to solar effects. This research should be placed in the framework of biorhythms which more and more, are shown to have an important effect on all earthly life. This point is still ignored by some technocrats who play with time as they see fit while giving fallacious arguments. However, this is not the place to prove this point.
FIGURE 8 ~ Variations of Ca after Dubrov
(6) Comments on Some Experiments ~
I will not go into the details of all the experiments performed; this would lead to tiresome repetitions. I would like to draw some lessons from the statistical data collected on tens of thousands of oats after hundreds of analyses of oats after hundreds of analyses on oat and oat seedlings. The information mentioned below is important, because it constitutes a base of preset references to verify if an analysis has been properly made. This is a point of the utmost importance for all researchers in order to avoid hasty and erroneous conclusions, some of which I noted in the research by students in national schools of agriculture or in public high schools which train superior technicians for agriculture. I noted the same trend among students as well as professors in schools of sciences in universities. I also saw so many engineers, considering themselves as specialists or experienced professionals, commit serious errors. These errors looked shocking to me as these people were in known territory, but apparently it was unknown, because they were prejudiced by previous experience.
I believe it is important that each set of seeds in a good stat of health and of approximately the same size. Each set should be prepared by the same operator. Unfortunately, in some schools several students prepare sets of seeds for the research which to be the subject of a dissertation or a thesis. Under these conditions, there are unacceptable differences between tests. Sometimes I noted weight variations reaching 20% and more, which biased the results. Each set should be weighted to 1/10 mg for control, as long as each set does not contain more than a few hundred seeds. Weighing should be done to the milligram when a few thousand seeds. The statistical average is then valid, barring bad luck. It is advisable to always avoid seeds of a common grade and to use selected seeds. Even if they first underwent mechanical selection. One must do this for several reasons:
(a) There are large differences between old species of black oats (some are still sown today, such as the Noire de Prieure for example) and a modern hybrid light seed. The differences can be as great as 100%. Often a species degenerates in less than 10 years. This was the case in the past for the black winter oats that we used. These seeds each weighted 44 mg in average. For the light Nuprime species hundreds of seeds showed an average individual weight of about 21-25 mg (depending on the origin).
(b) Black oats are winter grains. They grow well only if they are sown in the fall and only if they have spent the winter in the ground. The cold weather effect is absolutely necessary for a good start in the spring. It is called wintering. This phase in the enzymatic modification of molecular structures under the action of cold is seen as an absolute necessity, but the wintering is not yet well understood. Consequently, one should not forget to expose the seeds to artificial cold for several weeks when cultivating them in a laboratory. Too often, physicists neglect this step. On the other hand, spring oats and most light oats do not require wintering. Light oats can germinate at any time. Some precautions must be taken, such as varying the daily light exposure. There may be other cosmic effects, still unknown and little studied. Studies on this subject were made in Russia, Italy and Belgium. In France this research was mainly done by Prof Faussurier, Director of the Physics Laboratory at Faculte Catholique of Lyon. He used chromatographic techniques on metallic salt-impregnated paper in order to correlate color changes in the paper with cosmic phenomena which had been recorded by various observatories. There are still other effects, still unknown, which produce barely significant variations.
(c) In light all season oats, I stopped the research on the Panche de Roye species for two reasons: in this species the glume (also commonly called chaff) is well developed. It is necessary to eliminate it to avoid complicating the research after germination. Some agronomists maintain that the glume must be left because it participates in the germinating process. In my research, its effect on mineral balances and germination power always looked negligible.
In this variety there are two seeds in one envelope. Nine times out of ten, one is significantly smaller than the other. It is then necessary to open the glume to extract the larger seed, which complicates the operation. From the beginning this is why I concentrated my research on so-called naked or glumeless species, such as Nuprime which is cultivated in France and other countries mainly to make rolled oats. Later I used Flamingskone and Peniarth because I observed that Nuprime degenerated and was giving only 70% germination. One should use seeds sized and selected by a special section of INRA and with a minimum guaranteed ratio of 95%.
As a general rule, it is preferable to express analytical data in per unit values --- one seed, one plant. For comparative purposes one can indeed use equal weights and use the milligram or the gram as the unit of measure. However, when a seed does not germinate, germinates poorly or becomes moldy, the whole data for the seed is void. Therefore, the unit chosen should be the seed, except in experiments involving thousands of seeds. If one seed in a set of 100 does not germinate, the uncertainty is of 1%. Germinating ratios for the best seeds can only be guaranteed to 95%, leaving an excessive uncertainty of 5%. This is why I always insisted that sets prepared for cultivation always include several hundred seeds. How conceited on the part of researchers in Europe and America to be satisfied with sets of 20 or 30, sometimes less. In one experiment, for example, 6 seeds were left at the bottom of a test tube where they poisoned each other with the gases from their metabolism, causing a defective synthesis of growth hormones, etc. If a single seed out of 6 does not grow properly, a 16% uncertainty ensues. This is unacceptable. Unfortunately, these concepts of vegetal physiology are too often ignored by many physicists who only reason in the mathematical abstract. Many chemists specialized in inorganic analysis do not know biochemistry.
Personally, I always used sets of at least 50 seeds and several sets in parallel. Zundel often germinated batches of about 145 seeds each. He analyzed each set separately. he had two trays in operation for a total of approximately 300 seeds.
To avoid an uncertainty of up to 5% due to deficient seeds, it seems advisable to sue pregerminated seeds. To this effect, a number of seeds significantly higher than the number to be cultivated, is germinated. The seeds are placed between two sheets of filter paper of the so-called ashless quality. The seeds should not touch each other. The paper is soaked with twice distilled water continually; it is sprayed twice a day. Seeds germinate. After 4 days, the shoot (radicles, etc.) is a few millimeters long. Seedlings, which have germinated normally and are of comparable lengths, are then selected. One is reasonably sure to deal with seedlings of equal vigor. Each seedling is carefully transplanted in a seedling tray. An equal number of seedlings, as similar as possible, are selected, dried and analyzed for their respective contents in Ca, K, Mg, etc. An observation of the results shows that there s no appreciable difference in the weights of these minerals between fresh seeds and pregerminated seedlings. The synthesis of new minerals starts only after 5 or 6 days. It is therefore necessary not to overextend the pregermination period. After 15 days, for example, the difference in Ca is significant. This appears clearly in the curves we traced for calcium. Other cations and anions have also changed at the end of this period.
Hundreds of experiments showed that the relative contents of the major minerals in the seeds varies from one variety to the next. This is why some varieties are preferred when looking for specific elements. In the same species, variations may exceed 20% according to the variety. However, the composition spectrum is a characteristic of the species. In the same variety, relative variations between elements can reach approximately 5%. This is due to differences in the cultivated terrain, in soil composition, in the climate, in average seasonal temperature, in insulation, moisture, etc. This constant value of no more than ± 15% allows one to detect significant errors in the analyses. For example, with Nuprime seeds, I found the average seed weights of 22 mg (± 10%) according to the origin of the batch. The average Ca content was about 0.025 mg, or about 1/1000th of weight of the seed. With Peniarth seeds, the average Ca content per seed was 0.029 mg for an average seed weight of 28 mg, ± 1 mg. These measurements were made on thousands of seeds from the same crop. Some years I happened to note that the average Ca content for the same variety was lower by a few points than the 1/1000th of the weight of the fresh seed in Ca. This is due to climactic conditions during the year, or to the area of origin of the crops, or to intangible cosmic factors. This is of little importance.
On the contrary, if data differ by ± 15%, we can state that there was an error in the analysis, in calibration or in the preparation of the samples for the analysis, I have seen variations greater than 10%, which did not seem to worry the researcher.
Often it is interesting to take into account the normal K content and to compute the K/Ca ratio in the seed and in the seedling. For complementary information one may also look at K/Mg and Mg/Ca. This can lead to the detection of an error. In oats K/Ca is found to be close to 4.8 ± 0.5 with minor variations according to the species. This shows that the calibration probably is correct. However, comparisons with other analyses are indispensable, because errors in acid dilution may have occurred prior to injection into the analyzer. The K/Ca ration is not an absolute criterion of the validity of an analysis, but it often gives a good clue.
To illustrate this point: in an engineering school a student found an average Ca weight of 0.021 mg and an average K content of 0.078 mg per seed on 9 batches of Nuprime oats. This gave K/Ca of 3.72, which was a little too low. Elsewhere, several analyses of Peniarth seeds coming from the same stock gave average values respectively of 0.0273 mg for Ca and 0.1130 for K, or K/Ca = 4.1. This showed that the analyses of K and Ca were only marginally acceptable and that they had to be cross-checked, as ratios were found to vary with the species.
In Switzerland in 1974 a study made at a government laboratory by means of neutronic activation gave the following respective results for each of the 5 analyses: K/Ca= 4.5, 4.7, 4.6, 5.2 and 5.0, or an average of 4.8. It is a good average. However, the batches included only 6 seeds each and the accuracy of the analysis was only of 4%. The laboratory then made a correction for the seeds which had been poorly germinated. Computations were made on the base of 28.32 instead of 30. After correction the experiment showed an increase in Ca of 138%. In a French agricultural engineering school in 1973, the analysis of 9 batches of 30 seeds each (a slightly low number) of Nuprime oats showed average K and Ca of respectively 3.42 and 0.61 mg per seed, or a K/Ca ratio of 5.6. This was slightly too high a figure, perhaps because of an excessive K (calibrating error?) but in my judgment certainly because of an underestimation of Ca, due to an inappropriate measuring procedure for Ca. The measurements were made with a flame spectrometer. For divalent elements there is a risk that the number of atoms with no peripheral electrons is too small to yield reliable measurements. In a West German university, a study made by a student on Peniarth oats gave a Ca content of 29.5 micrograms, a figure which seems too high for seeds with an average unit weight of 21.36 mg. It is almost certain that an error was made on the K content, given as 118.36 micrograms. This leads to a K/Ca ratio of 4.0, which is too low, but shows that errors on K and Ca can compensate each other. Therefore, it is necessary to do several cross-checks. I will not mention any figures for a study made in a university in the USA, because the results are certainly wrong. Though one might have reservations on x-ray fluorescence measuring techniques with K and Ca, this time the error came from a mistake in the growing procedure. Cultivation was made under artificial lighting with a lamp giving a completely inappropriate spectrum. This resulted in deficient photosynthesis.
A set of studies on 10 batches of Peniarth oats coming from the same stock gave 3.9 for K/Ca. Several months later 8 batches from the same stock led also to K/Ca = 3.9, showing first that there had been no drift in the composition and second that the concentrations were a little too low for this species. Usually the average figure gravitates around 4.8 ±10%. An error could have happened either in the growing procedure or in the analysis. Once again we see that errors compensate for each other, which makes cross-checks necessary.
An easy cross-check of the data in a study is provided by the seedlings dry weight. It must be greater than the seeds dry weight because with good growth, there is production of carbohydrates (starch, cellulose and other glucids, lipids and parotids) due to seedling growth. This is due to the photosynthesis metabolism. The weight increase was observed and measured as early as the 17th century. Nevertheless, I noted that some French and some foreign researchers mentioned figures showing no increase in the weight of dry material. Some even showed a decrease. This points to an error in the growing procedure. This did not even catch the attention of these researchers. I will come back later to the importance of good photosynthesis. I would like to say here that only dry weights can be validly considered. Long discourse on the necessity of stabilizing the weights according to the humidity are without merit, because moisture in the laboratory varies with the season, etc., and comparisons between successive experiments are not valid. Dessication standards should be set.
Even the weight of ashes is found to be higher for seedlings than it is for seeds in most cases; this may be attributed to methods of analysis inappropriate for these types of studies. For example, in chemical analyses of ashes, losses of phosphorus and sulfur, among others, during firing are often forgotten. Some compounds become volatile at fairly low temperatures, sometimes as low as 200° C, which is insufficient to obtain good ashes. One forgets also that calcium sulfates and phosphates in some cases are insoluble in the acid used and they may be formed during seedling growth. In these cases there is an excess of ashes, but the amount of calcium is underestimated by the quantity which remains insoluble. This shows that many chemists do not know their trade. If hydrochloric acid, often used, does not give satisfaction, even if it has been heated, then other acids, such as sullfo-nitro-perhydric acid or hydroperchloric acid should be used for the tests. However, we should not forget that these acids are potentially explosive and they are difficult to handle. Usually hydrochloric acid will do, but it is a good precaution to test the potential margin of error to see if it is acceptable. Physical analytical techniques applied to dried, instead of incinerated, powders can be used to circumvent these difficulties. However, with some techniques involving the use of physics instruments, solution is still part of the procedure and the dissolving power should be taken into consideration. No such problem occurs with spectrometers when the powder is sparked.
Some of the figures mentioned in various studies are so obviously wrong! Such is the case in a study made in a German university on 192 Flamingskrone oat seeds and 190 seedlings. After 49 days of growth the following data was reported:
Average dry weight of each seed: 21.77 mg, and
Average dry weight of each seedling: 32.67 mg.Data reproduced without comments.
There is certainly an error. Furthermore, the contents mentioned for Ca, mg and K are questionable. All these figures are smaller than corresponding figures for the seeds, though the total weight increased. The Mg/Ca ration is completely abnormal. Other cross-checks show that the figure for Mg is much too low, and that the figure for Ca is too high. Evidently this student did not master the analytical technique he was using. In an American university we saw similar results, although the supervising professor did not make any comments. Every result should be cross-checked.
In another university department the following values were quoted in micrograms, per Peniarth seed:
K = 11.4; Mg = 33.7; Ca = 29.5, which gives:
K/Ca = 4.0; K/Mg = 3.5; Mg/Ca = 1.1.K/Ca is almost normal. This might lead one to believe that K and Ca were fairly accurately measured. However, if:
K/Ca = 4 and K/Mg = 3.5, it is impossible to have Mg/Ca = 1.1
A French researcher told me that he found the following relations:
K = 0.113; Mg = -.030; Ca = 0.027
K/Ca = 4.1 (about normal); K/Mg = 3.6, and Mg/Ca = 1.1.These values are very close to those quoted previously and show an error of the same order of magnitude in Mg.
In official tables for an unidentified species and following analyses by purely chemical means, K/Ca is given as 7.03, which is too high. This shows that the value measured for Ca was too low, because Mg/Ca is given as 2.5. If Ca is too low, then Mg is also too low, as the K/Mg is given as equal to 3. At the minimum the chemical analysis of Ca and Mg should be checked. It is likely that Ca and Mg were not properly separated (probably with oxalate?). This would explain the errors in the three reports mentioned.
Experiments correctly performed show that K + Mg + Ca + Na is approximately constant for a given species and no additional fertilizing elements are brought in. Whatever the species, one can favor any cation by selection. Na varies a great deal in relative value (often 50-60%), but we neglected it because it is very small in absolute value in vegetals. Therefore, it is permissible to only measure and compare K + Mg +Ca. Many experiments showed that Mg does not vary significantly in oats. For oats it is therefore sufficient to compare only the values of K + Ca to detect gross errors in tests or analyses. I also neglect Si, which shows small variations in relative value. However, these variations are often interesting to study, as observed by Vauquelin in 1799. Please refer to my book of 1975.
By rounding off the numbers in the previous example, one obtains K + Ca = 0.14 mg in the seeds as in the seedlings. This leads one to infer that the analyses are correct in both cases (there is a small error for Mg). On the other hand, I noted an error in a study made in a foreign university in which K + Ca was given as 0.154 mg for seeds and only 0.145 mg for seedlings. In this test there was less Mg in the seedlings as well, and the researcher did not notice that all the elements were in smaller quantities, although the dry weight had increased by approximately 50%. With such comments one can appreciate the validity of experiments. If Mg is also measured, we will note again that the three ratios, K/Ca, K/Mg and Mg/Ca help to determine which cation was in error. I shall mention again that I gave the following figures for the Flamingskrone variety:
Seed: 0.1130 K + 0.0273 Ca = 0.1403 Total
Seedling: 0.0799 K + 0.0590 Ca = 0.1389 TotalBy rounding off to the third decimal:
In the seed the total is 0.140
In the seedling the total is 0.139,which is a close match. We can refer to numerous and various cross-check performed on tens of thousands of seeds of various species. However, in seedlings, the dispersion of the measurement is often considerable due to differences in growing procedures: light spectrum through various materials, etc. In any case K + Ca should remain nearly the same in the seed and in the seedling. If not, one should try to determine the cause of the error, if possible.
(7) Reservations on Some Analytical Techniques ~
Several times I pointed out that a physical analysis technique, requiring the most modern and sophisticated instrumentation, could not always be accepted without reservations. One must always cross-check the results by completely different techniques. For example, I mentioned the case of an error due to a wrong dilution during an analysis by atomic absorption spectrophotometry. For comparative purposes, the gas pressure fed from a liquid acetylene supply, the gas flow to the Bunsen burner, etc., should be recorded. So should the lanthanum concentration in the buffer solution, which prevents interferences by some unwanted elements. I also pointed to some sources of error in electronic transducers connected to a computer. Each class of instruments has limits. I do not question how the figures were obtained, however they are not always reliable.
(8) One Example With X-Ray Fluorescence ~
There are crazes, fashions and also habits peculiar to the laboratory or to the operator. In Figure 9, I show an example of a curve obtained by x-ray fluorescence from dry ground oats in the nuclear physics laboratory of an American university. It was taken from recording made in 1976 of the simultaneous measurements of K and Ca. The continuous curve corresponds to the actual measurement. It seems that, starting with an atomic mass of 41, the K peak has a steeper slope. Studies have shown that this is due to the Ca curve which contributes to the sharp rise in the K curve. The K peak is not a line. It spreads widely and masks the curve for Ca. Computation factors fed into the computer are somewhat arbitrary, because they may vary according to the material analyzed. For this reason the final result may be right or wrong. A cross-check by a different technique is necessary, no matter how carefully the calibration was performed (as the zero adjustment of a pH meter, in the case of a relatively simple instrument). The values for the K peak and for the Ca peak (dotted line) are good. A similar analysis was performed after the germination of the seeds. For the species tested, it showed an increase in Ca of 125% corresponding to an equivalent decrease in K, which confirmed the correlation between the two elements.
FIGURE 9 ~ Analysis of K and Ca by X-Ray Fluorescence
Additional Information On Physical Phytochemistry During Germination ~
Too many physics specialists are tempted to simplify vegetal biology processes and they select germination procedures in which normal reactions cannot develop. This results in distorted data. I do not intend to give a detailed study of this topic, but I want to describe a few steps in the preliminary study. This study is essential if valid experiments are to be achieved.
(1) Summary of the Germinating Phases of Grain Seeds ~
When the seed is exposed to a temperature of 15-20° C and to the proper moisture (close to 100%), the embryo produces which migrate toward an external layer of the seed, located under the tegument and made of aleurone cells. There the hormones cause the formation of various types of hydrolytic enzymes which circulate between the cells in the core of the seed (endosperm). These cells constitute the reserves of the seed. The enzymes destroy the walls of the cells and transform starch and proteins. Under the action of diastase, starch is transformed into various sugars.
An enzyme cannot act if the cell structures constitute an impermeable barrier and prevent the enzyme from reaching the substances it is supposed to transform. This is why some enzymes must first transform the walls of the grains of starch. There is, therefore, a programmed sequence of enzymatic actions. The product of a reaction with an enzyme constitutes the substrate for the next reaction, which includes a spatial modification of the molecular structure. Successive enzymes are prepared and they become available at the right time, so that the effective section of interaction with cosmic neutrinos is continuously changing.
In the fresh seed, the external envelope, or tegument, provides a mechanical protection for the endosperm, which constitutes the reserve of food. The embryo is located at one end, and is also protected by the tegument, from which the roots and leaves will sprout. As early as the second day, the embryo synthesizes a growth hormone, gibberellin. This can only be done in the presence of water and under favorable temperature conditions. It acts as some sort of a messenger for RNA which reaches the aleurone layer in a few days (1-2 days). There it produces the synthesis of hydrolytic enzymes, which will transform the structure of the grains of reserve proteins. These proteins include avenin (in oats) in addition to structural proteins, the glutelins. The endosperm cell walls are mainly constituted of beta-glucane chains, which are glucose polymers, close to cellulose, linked by very short peptid chains.
One of the effects of seedling growth is the decomposition of these various constituent elements into small molecules. A few hours after the seed is moistened, the peptid links between the beta-glucane elements are broken by an enzyme, carboxypeptidase. The degrading process develops step by step. A whole series of enzymes act specifically on the internal links as soon as they are in contact with the products decomposed by the previous enzyme, and they produce various sugars. During that time, other still relatively unknown enzymes decompose the pentosanes, breaking the walls of the starch grains. Once the cellular walls are broken, other proteins interact with starch, which is converted into glucose. During this time, the gibberellin migrates to the opposite end of the seed (the distal area) over a period of 10 days, transforming almost all the aleurone cells at the back of the seed. By that time almost all of the starch has been structurally transformed. The transformations are by then macroscopically significant, to the point of making variations in the seed chemical composition measurable. We published curves showing the abrupt changes which happens after about 10 days of germination.
After two days, the tip of the embryo starts to emerge from the tegument. On the average, the radicle length is 2-3 cm after 4 days. The embryo has eaten close to one-half of the starch by this time. After 8 days, the radicle has 608 offshoots and almost all the starch has disappeared. Enzyme activity is then fully accelerating and analyses show very significant biological transmutations. It is at this time that the photosynthesis contributes its effects as leaves are already well developed. This results in a sharp rise in the calcium content shown in our curves around 8-10 days. Analyses made at short time intervals show that seed compounds are not appreciably modified until the fourth day. For the purpose of comparing Ca contents, after 28 days for example, it is legitimate to select the seeds which sprout well after 4 days of pregermination. It is understood that no Ca should be brought from the outside during that time.
FIGURE 10 ~ Grains of Cereal in Germination
(2) Average Curve of the Increase in Calcium of Oats After Germination ~
To my knowledge, about 60 experiments were performed over the last 10 years on the germination of oats to study the increase in calcium in hydroponic cultures. This was done over a period of several weeks without external addition of Ca.
These experiments required over 400 analyses by different physical or chemical techniques. These analyses were made on tens of thousands of seeds or seedlings. The increase in Ca is measured by comparison after analyses of a seedling and of a seed similar to the one from which the seedling originated. This is to say that the phenomenon was studied for a long time and that it is firmly established.
It seems of interest together the results from approximately 40 experiments especially well performed on light hybrid species only. Their seeds are small and generally more homogeneous than the big black seeds produced by old varieties. Nuprime, Flamingskrone and Peniarth varieties were primarily used in these experiments. The seeds weighed approximately 20-30 mg according to the species and the batch. They were calibrated to 1 mg after selection in batches of 100-300 seeds. Some analyses involved batches of more than 1000 seeds. The units retained here for comparative purposes are the seed and the seedling grown from a seed, because they represent the minimum biologic quantum of enzymatic action. If a seed does not germinate, the whole weight of the seed remains inactive. However, seed weights should always be noted to allow comparisons between batches of various origins and varieties. In the light hybrid varieties considered, the Ca content represents approximately 1/1000th of the weight of the seed. This figure is based on tens of thousands of seeds. A fresh seed with an average weight of 25 mg will have roughly a calcium content of 0.025 mg.
Figure 11 summarizes the data obtained after about 40 experiments. This does not mean that each point of the curve corresponds to an average of 40 analyses (by different techniques and on several varieties). Most experiments involve only the analysis of Ca in non-germinated seeds, and then on the seedlings grown from similar seeds and harvested after 4, 5 or 6 weeks. Experiments, in which seedlings were sampled after 2-3 weeks, are few; the majority of the data is related to seedlings cultivated over 4-7 week periods. However, data collected either during a short experiment or sampled after a few weeks from a batch left under culture, are compatible and they allow one to draw the curve in the appendix.
This does not mean necessarily that every figure found for Ca in a new experiment or a given duration will be located on this curve. It is an average for several varieties obtained by means of various analytical techniques. The dispersion inherent to biological batches of different vigor should always be taken into consideration. Nevertheless, the effects of all these elements should not cause a variation greater than 15-20% from this curve even if they are cumulative. Hence, this curve can be used as a guide in this type of research, to detect a major error in the procedure or in the analysis. We established it for this purpose. It is a piece of information and it gives an indication about the magnitude of the phenomena involved in the production of Ca under the effect of growth hormones. One should note the asymptotic trend of the curve after 2 months of growth (these spring oats are normally harvested 4-5 months after sowing). The growth metabolism is only active during half the life of the plant. Growth hormones (gibberellin, auxins) synthesize the enzymes for the plant anabolism during germination. The curve shows that the Ca formation is quantitatively linked to the activity of growth hormones until exhaustion at the point of maturity (1)
[(1) The diminishing in growth of Ca production may come from Ca saturation in the plant which, in turn, affects the enzyme. The saturation is reached after 8 weeks with a K/Ca ratio approximately equal 1.5. In the seeds the K/Ca ratio is approximately 4.5. The transmutation of K into Ca stops when the two elements are about equivalent in weight. That would be at the time of maturity, when the growth hormones effects have ended.
FIGURE 11 ~ Augmentation of Ca in Oats
However, one should not infer that the production of enzymes alone can explain the energy balance for the transmutation which leads to the production of Ca. The enzyme production constitutes an essential element because a seed which does not germinate, does not produce any biological transmutation. The question of energy does not have a place here. I only wished to present an objective document on the data obtained as reference for future research.
Chapter 4Photosynthesis
In one of my early books I mentioned a 17th century experiment made a Flemish physician, Jean-Baptiste Helmont, in which he planted a 5 lb willow tree in a planter containing 200 lb of soil. The planter was covered with a lid with two holes, one for the trunk and the other for watering the tree. The purpose of the lid was to prevent any changes in the weight of the soil by dust from the atmosphere. After 5 years the tree was uprooted and weighed. It weighed 164 lb. However, the weight of the soil had only decreased by 2 oz. Helmont did not find any satisfactory explanation for this observation. Photosynthesis was then unknown.
For a long time the study of the phenomenon remained sketchy. From time to time crazy results were quoted in quasi-official documents, without attempted justification. For example, it was stated without comment that soils were improved by bamboo trees (1969). Oureschi, Yadar and Prakash observed that the species Bambusa Tulda gave back to the soil more calcium than it took. Other bamboo species, such as Nechouzeana Dulloa and Oxytenanthera Nigrociliata, "give back more magnesium" --- from Nature and Resources No. 4, p. 15 (1975), UNESCO Publications.
But it was only during the 70s that the mechanism of photosynthesis was studied more thoroughly. It was finally understood why monocotyledons exposed to atmospheric carbon dioxide gave compounds which differed from compounds found in dicotyledons, C4 compounds in the first case versus C3 in the second. This explained differences in behavior and yield among various families. Some plants growing on salty soils follow the C4 cycle due to a specific effect of Na. This is also true of many weeds. They take more carbon from the ambient air, and therefore need less fertilizer than plants with a C3 cycle. In addition, they produce more dry matter and consume less water. We will study the role of photosynthesis more closely, a subject still ignored too often by many agronomists, because it was not taught to them.
(1) Effects of Artificial Lighting in the Photosynthesis, for the Study of Transmutations by Cereal Plants ~
Many experiments were performed in various countries at various times to study the transmutation of particular elements during plant germination.
Usually they were performed under natural light. With the development of this research, a problem related to the use of artificial light arose. It was used in particular for the studies on the germination of cereal grains, especially of oat and wheat.
These grains, as well as barley and rye, are said to have long day cycles. This means that they grow in daylight and reach their maturity shortly after the longest days in the year. Ideally they need 13-15 hours of daylight in French latitudes, 16 hours in more northern latitudes.
For this reason, the research on transmutations, which are produced at the peak of the enzymatic activity (strong growth of the plant) is distorted when laboratory experiments are run outside the spring season. Transmutations are indeed observed during germination in sunlight, at different periods in the year, but they are not quantitatively equivalent.
It became clear then that artificial lighting would be beneficial, even in spring, in a room insufficiently exposed to daylight. It would also be useful for studies made at other periods of the year to extend the action of sunlight. It would enable the experimenter to have an artificial day of controllable length, so as to always operate under optimal conditions for photosynthesis in all locations and seasons.
(a) Photosynthesis ~
We are not trying to summarize here what photosynthesis is. We are restricting this word to mean the action of solar rays on the chlorophyll metabolism. This does not encompass the whole of photobiology. We will not discuss more specific subdivisions such as phototropism, which manifests itself by an orientation of the pant toward light. As a point of interest for what follows, we will note that blue rays are most active for phototropism. Oat plants lit laterally with blue light bend toward light. As a point of interest for what follows, we will note that blue rays are most active for phototropism. Oat plants lit laterally with blue light bend toward the blue light source. There is an antagonistic effect with longer wavelengths. If the other side of the oat seedling is lit with a green or yellow-green light, the seedling grows vertically. We will not touch either on what is called photoperiodism, or the influence of alternating periods of light and darkness. We shall recall that the inverse reactions of photosynthesis are produced in darkness by the plant. Photosynthesis achieves the global effect of taking carbon dioxide from ambient air, fixing carbon in various organic compounds (carbohydrates) and ejecting the oxygen generated from the water generated from the water supplied to the plant. In darkness, the plant takes oxygen from ambient air and ejects carbon dioxide. This is respiration, which only becomes important in darkness, because there is no photosynthesis. Furthermore, we will not discriminate between the optical spectrum of photosynthesis and the sensitivity of chlorophyll to various wavelengths of the solar spectrum. The curves for these two phenomena are very close, but they do not coincide, because photosynthesis includes additional reactions beyond the metabolism of chlorophyll.
It is essential to remember tat our eye and plants have much different sensitivities to various wavelengths of the solar spectrum. It is not sunlight, as we see it, which should be used as a criterion in looking for the artificial light source closest to sunlight. We must find out to which parts of the solar spectrum chlorophyll and the whole plant react best. A difficulty stems from the fact that our measuring instruments, luxmeters for example, are calibrated from the effects on our eye. Brilliance and lighting units are defined by optical effects. It is the eye which is the final standard of measurement. In fact, there are two completely different aspects. This appears obvious in Figure 12a where we have drawn the curves of maximum sensitivity to wavelengths for the eye, and for photosynthesis.
FIGURE 12 ~ Lamp Spectra
Wavelength is expressed in nanometers (nm) or billionths of a meter. This is the unit most commonly used today, because it conforms to the metric system. It is the thousandth part of a micrometer (or a micron). The millimicron and the nanometer are equivalent, the second being more common in spectrometry. Some people use the angstrom unit, which does not belong to the metric system and which for this reason is being progressively abandoned. One anstrom is equal to 10 nanometers. People using microns will divide by 1000 the figures quoted here in nm.
Visible light extends in principle from 400 to 700 nm. These are average limits. In fact, some people can see down in the violet as low as 380 nm; others see up in the red as high as 760 nm (sometimes up to 800 nm). By convention, the near infrared (IR) encompasses the 750 (0r 800) to 1200 nm band. The far IR covers approximately the band up to 3000 nm. Beyond this wavelength the action of solar radiations is negligible and its effects on photosynthesis is not considered. The longer the wavelength, the lower the frequency associated with the wave. The photons with the most energy correspond to high frequencies, hence to sorter wavelengths; they are the photons in the blue, violet and ultraviolet (UV). Rays with wavelengths shorter than 380-400 nm are rated as UV.
The solar spectrum includes UV with a minimum wavelength of 390 nm (or 288). Shorter wavelengths are stopped by the ozone layer surrounding the earth. Life on earth is only possible because of the filtering out of the rays of shorter wavelengths. In fact, shorter UVs are used for their bactericidal effects. They are life destroyers (the energy of their photons is such that they decompose living molecules).
For photosynthesis one should not look for lamps with rays shorter than 290 nm. Even in photosynthesis no action is observed from rays with wavelengths shorter than 380 nm. In fact in nature, in the open country, there are rays between 290 nm and 380 nm. What is their purpose for the plants? They may act on pigments and on protovitamins, in animals as swell as in plants. There are few, quantitatively, and their energy seems to be weak. Their study is indeed difficult. In order to do this, one must find solid substances to make prisms transparent to these wavelengths and opaque to others. The molecules of monochromatic substances should not be ionized by the energy of the photons in this band. Such materials are delicate. Therefore, we do not know what nature does with the rays in the 290-380 nm wave band, perhaps because their study presents too many difficulties. We are noting this, but we will not come back to it, because the present document is only related to the aspects of photosynthesis which we can understand with the help of todays instrumentation.
In fact, it seems that photosynthesis is altered by wavelengths shorter than 380 nm when they are of high intensity. This is the case of the 365.4 nm mercury ray. It penetrates through quartz, but it can be stopped by glasses of various compositions. For the purpose of photosynthesis, glass filtering UVs below 375 nm should be used in mercury discharge fluorescent lamps (or tubes) when fluorescence is produced by the internal coating. When the arc is produced between two electrodes in mercury vapor, the discharge tube must be made of quartz so it does not stop the UVs, which will trigger fluorescence in the special powders coating the inside of the light fixtures external glass surface. The glass shields the plants from UV rays (below 375 nm), but it does not stop fluxes in the 375-400 nm wave bands which are essential for some phases of photosynthesis. The intense 253.7 nm ray is completely stopped by the glass and it is used for the excitation of fluorescence.
On the other hand, IR rays have weak energies. Their energy is essentially thermal. It is sufficient that the glass remains transparent to rays up to about 1000 nm. Glass is opaque to wavelengths in the far IR, which explains its use for the greenhouse effect. The near IR penetrates inside, and the ground reemits a heat of a longer wavelength which cannot get out.
The radiation of red and IR rays up to 780 nm controls the opening of the stomata of the leaf, hence its respiration and its transpiration. Beyond this point, a counter effect is started. Leaf moisture loss ad gas loss should not become excessive. Beyond 780 nm, photon energy is low; this is why the curve of photosynthesis intensity remains close to the horizontal axis (see curves in Figures 12a and 12c). The thermal agitation due to red and longer wavelengths excites electrons in the atoms of the organic material of the leaf. However, the energy is too low to permit the extraction of the electrons which have moved to less stable orbits. Electrons do not move from atom to atom as they would under the stronger energetic effect of blue and shorter wavelengths. This constitutes the chemical effect of blue, violet and UV rays on the molecules prepared by the thermal effect of red rays. It can be seen from the curves in Figures 12a and 12c that the peak of energy is higher on the side of short wavelengths than it is on the side of long wavelengths. By contrast intermediary wavelengths in the green and yellow have no significant influence on photosynthesis. On the contrary, under artificial light in an apartment, for example. Our eye is most sensitive to yellow. Consequently, lamps designed for industrial or domestic lighting should not be used in phyto-optics. We note this point, which is sometimes forgotten.
(b) Characteristics of Some Lamps ~
We will not list the tens of lamp types used for artificial lighting in agriculture, horticulture, etc. They are described in many published works. We used a wide selection of lamps, incandescent, discharge, fluorescent and mixed, because not all plants are sensitive to the same wavelengths. This fact is well known and it is used in the design of commercial products. Most often it is known only empirically and the related information is marred by many errors. We evidenced this during experiments on the transmutations observed on particular elements during the germination and the growth of cereal plants.
At the beginning of the 60s some agronomists recommended lamps of the warm white type or sometimes warm white deluxe. I believe this is a mistake. Figure 12b shows a diagram of the characteristic emissivity for these lamps. The rectangles show the primary characteristic rays of excitation of the fluorescence. The continuous curves characterize the spectrum outside the lamp. It is obvious that these lamps are very deficient in the blue-violet and too strong in the hot colors, the reds. We saw that red has an effect on the control of transcription, hence on gas exchanges. Transpiration depends on the energy in the blue for about 40% and on the energy in the red for 60%. However, plant growth and development and the chemical reactions which cause the synthesis of the organic compounds constituting the plant, are only active in the blue-violet and the UV. Growth hormones (auxins, etc.) are also synthesized under their action. We are not surprised that phytobiologists, under their action. We were not surprised that phytobiologists, after following erroneous procedures of 15 years ago, got underdeveloped seedlings. Their plants grew only a few centimeters after several weeks as seedlings of 25-30 cm height are obtained under optimum lighting in the same time span for wheat as well as for oat. I have no base of comparison for other grains. Without blue rays, wheat assimilates nitrogen and potassium with difficulty. Contrary to what happens under normal cultivation, the plant cannot assimilate K correctly, therefore no transmutation of K into Ca can take place. The circulation of water and mineral salts inside the plant is normally made because of the long wavelengths. It is only a transfer, which does not involve any physical or chemical transformations. These are only triggered by wavelengths in the blue through UV band.
The color temperature of the warm white lamp is of 3,500 K (or color 29 on the Phillips scale, as compared to 34 for the Warm White Deluxe. This figure of 34 corresponds to a radiated power of 7.1 watts, as the lamp of color 29, which radiates in the red, has a peak of 8.1 W.
The radiant flux is greatest for the Daylight Lamp, but its flux in the red is too low. Its flux in the blue is significantly higher. It is felt that it gives objects a bluish cast. This is why these lamps were replaced with lamps which gave a warmer light (more red, less blue). These lamps have a radiating energy in the blue which is significantly too low for photosynthesis. They are lamps for home lighting. We will note that these lamps strongly radiate in the intermediary spectrum, which is adapted to our sensitivity and appropriate for interior lighting. However, it is a waste for photosynthesis.
(c) A Well Designed Lamp ~
One can change the shape of the fluorescence curve by changing the nature of the powder coating the inside of the discharge lamps. This can also be done by changing the nature of the rays triggering fluorescence in the compound coating the inside wall of the quartz discharge tube.
Iridium radiates in the 411-450 nm band, hence in the blue. There is even a very weak ray of 380 nm in the UV. There are also two important rays around 680 and 690 nm. Two other elements are also used to trigger fluorescence: thallium which radiates at 534 nm and sodium at 588 nm. However, their intensity in the yellow must be corrected with too many blue and red wavelengths to be practical. These lamps are now of little use. Magnesium arseniate, fluorogermanate and manganese compounds were also used for the fluorescent coating. Iodine gives an intense ray at 577 nm, at the limit of the yellow and green. Because of this, it is used in large lamp fixtures for public lighting and automobile headlights. However, iodine has other useful properties. It also gives a ray in the UV and it is added in some fluorescent lamps as such, or as metallic iodides. Unfortunately, almost all the known spectra are not appropriate for phyto-cultivation, at least in studies on elementary transmutations. These transmutations cannot happen with spectra of the types represented in Figure 12b. I shall not comment on the Osram Power Star lamp used by Zundel, as I had no experience with it.
In Figure 12c, I am showing the spectrum of a lamp specially designed by Societe des Lampes Claude in France. For over 15 years lamps of the types in 12b proved inadequate. Societe Claude markets lamps rated to 250 W and to 400 W. I was gratified that they lent me units of the 250 W model giving 12,000 lumens. Lets not forget the relativity of the light flux concept for the eye. A close correlation between lamp and photosynthesis spectra is the only consideration which counts. In this oblong lamp, electrical discharges are produced in Hg at low pressure between electrodes inside a quartz tube. The tube is closed and it can be mounted in a horizontal position. It is surrounded by a surface made of special glass and internally coated with a fluorescent salt. Various compounds were tested such as a ytrrium vanadiate doped with europium; other more common compounds, such as phosphates, aluminates, etc., could be used.
Rays radiated by the discharge tube, such as the 253.7 nm Hg ray, trigger the fluorescence in the coating. Due to the filtering effect of the glass, the 404.7nm and other rays of longer wavelengths are the only ones radiating outside the lamp. The spectrum distribution is shown in Figure 12c. It is seen that two Hg rays are located toward the end of green and in the yellow. They have little positive effect on photosynthesis and no negative effects in general. Their effect is to give a light which is relatively white to the human eye, a useful point in greenhouses. There, flower and leaf colors should look the same as under solar light, so that the general state of health of the plants may be gauged in one glance.
The Phytoclaude 400 W model lamp, which was lent to us, gave the following radiated flux in watts for various bands:
Wavelengths in nm: 400-450 ~ 450-600 ~ 600-700 ~ Total: 400-700
Flux in watts: 10.2 ~ 2.29 ~ 0.92 ~ Total: 13.93One clearly sees the considerable advantage presented by this lamp: approximately 10/13 of the energy transformed in light is in the 400-450 nm band, which is the active part of the spectrum for photosynthesis. The Warm White Deluxe lamp radiates only 1.7/13.4 of its energy in the 400-510 nm band. In fact, the 520 nm wavelength is already in the green, hence has little effect on the photosynthesis. The total radiated energy is not a sufficient criterion; the important point is the band distribution. Even if the band is extended to 510 nm, the Warm White Deluxe lamp gives 6 times less energy than the Phytoclaude lamp gives in the blue-violet with similar total radiated energies (13.4 W vs 13.7 for the Phytoclaude lamp).
(d) Cultivation in a Closed Chamber with Carbon Dioxide ~
Commercial cultivation of flowers and some vegetables under phyto-lighting is usually done in large greenhouses. Laboratory studies are often made in small rooms and even in small cabinets designed for this purpose. Sometimes they are made in the corner of a laboratory under a plastic shelter which lets light in but stops dust. In all cases the plants are placed in an atmosphere renewed naturally or by mechanical means.
For more precise studies, one can use closed chambers maintained at a slightly positive pressure (3 mm W.G. for example) by a filtered air supply. Chambers normally include a removable panel to allow the handling of the equipment and seedlings. Panels are fixed in place by gaskets and screws. They are sometimes made of polymethylmetacrylate (plexiglass, for example) transparent to the right bands of the solar spectrum. These chambers can be exposed to solar light or to phyto-lamps. Even better results are achieved with Altuglas.
When extra precautions to prevent any external contamination are deemed necessary, seedling trays are supplied with air by an electric centrifugal lamp delivering a positive pressure. The air passes through a filter which retains practically all the dust, as we verified. Sometimes even more stringent precautions may seem necessary. Lets take the case of the study calcium variations during the germination of grains. One must operate with water and air absolutely devoid of Ca variations during the germination of grains. One must operate with water and air absolutely devoid of calcium to obtain an accurate Ca balance before and after germination. Sometimes the Ca was measured by neutron activation techniques in the seeds. Unfortunately, seeds exposed to non-lethal doses of neutron radiations were sometimes irradiated for the experiments. This was a mistake. Seedlings are watered with twice distilled water, free of measurable Ca. After several weeks the plants are analyzed by various methods. To prevent any dust from entering the plant chamber the supply air is bubbled through a flask partially filled with an acid solution (30 ml HCl to 750 ml of twice distilled water in one-liter flask). The air is then pushed into the hydroponic cultivation chamber. Any lime particulate in suspension is dissolved in the acid solution. With the positive pressure, no external air leakage into the chamber can occur, even if a gasket is defective. Any leakage would be detected with the water gauge at the air intake.
With these precautions, numerous studies on oats showed that there was a production of calcium by the seedlings during their growth.
Additional precautions were taken for studies on oat and wheat in particular concerning the purification of the air supply. Behind the flask with the acid solution were added two flasks containing a NaHCO3 solution and one flask containing twice distilled water for a final scrubbing. The role of the basic solution was to neutralize any acid carry-over without trapping the CO2 contained in the air supply. These represent the most elaborate precautions which can be imagined. This was necessary in order to counter prejudiced objections, which ascribe all increases in calcium to an external source. Even with such a wealth of precautions, there is an increase in Ca which can exceed 100% of the total Ca contained in the seeds germinated.
Tested were made on wheat in a controlled atmosphere with a CO2 concentration of 0.08% and a lighting 4,000 lux. With 10,000 lux it was possible to raise the concentration of CO2 up to 0.013%. Under intense sunlight it is possible to reach 70,000-80,000 lux, but the CO2 concentration cannot be increased in the same proportion. There is a saturation phenomenon in the plant for various reasons. Principally because other functions must follow: water and mineral salt circulation, gas exchanges, etc. For wheat, the optimum concentration seems to be 0.5-0.6% or approximately 15 times as much as in ambient air, and it produces an assimilation about ten times higher than normal in the atmosphere. We shall remember that the lux measurement is of little value as it refers to the human eye. Studies were made up to 20,000-30,000 lux and even more, but this is not economically feasible.
On the other hand, good results were obtained on wheat in a controlled atmosphere including 0.14% CO2, which is four times the natural concentration. These results were obtained in a glass chamber in sunlight without additional phytolighting. More straw, heads and grain were obtained and the heads were larger. This shows that the development of grains is limited by the relative scarcity of CO2 in the atmosphere. In a controlled atmosphere CO2 concentrations 4 times greater than in ambient air can be used. We shall note that most published data related to this subject are wrong because they are based on Calvins cycle. We shall comeback to this point.
One barley species shows great resilience to intense irradiation by a light source of 460 nm. In this band, 10,000 kiloergs/cm2 are required to stop flower formation. By contrast, it takes only a much smaller amount of energy (about 1,000 kiloergs/cm2) at 480 nm to stop the formation of soybean flowers. Consequently, in the open, barley blooms during long days and soybean plants bloom during shorter days, when the intensity of the sun has subsided. Usable energies in the read are always low. Barley needs only 300 kiloergs/cm2 at 700 nm. The figure is the same for soybeans. Barley and soybeans show similar effects to green light. Grains use 10 times as much energy in the blue as the legumes. This must be taken into account when using artificial light. There is no general rule; each plant species should be studied separately.
(e) A Few Additional Comments on Photosynthesis ~
I will deal here with a subject which, to my knowledge, has not been presented in any published work. Our knowledge of the photosynthesis phenomena is incomplete and agricultural engineers should hold reservations regarding some so-called authoritative statements. We observe some results, but we cannot explain the complete sequence of reactions which lead to them.
The most common mechanism for explaining photosynthesis is the Calvin cycle. This cycle was adopted with much enthusiasm and too little discernment, because there was no acceptable explanation previously. Calvin started from existing intermediary organic compounds, which were already isolated. Everything Calvin surmised from there on seemed to link up logically. However, he neglected energy balances. Many experiments with similar results seemed to show that 8 photons were required for each CO2 molecule in the air in order that the plant could synthesize organic compounds. But what plant? Conclusions true for a particular plant were incorrectly extrapolated to other plants. For these, the Calvin cycle required a much higher number of photons. Consequently this cycle, adopted with enthusiasm by a majority of the people, was the subject of justified objections. The cycle was modified several times and Calvins name is now followed by the names of the main researchers who added to it.
Despite all these additions, some organic molecules formed by photosynthesis in certain plants could not be explained by the Calvin cycle. Following a very human and common tendency, studies of photosynthesis had been conducted mainly on a single plant, spinach, which supplies green material in large quantities and almost all the time. It was a fashionable plant. The results were extrapolated to every plant. There was one photosynthesis mechanism. It was concluded that chlorophyll (a and b in particular) was the same everywhere, and consequently that chemical reactions had to be identical in every case.
Experience contradicted this simplification. It was observed that the Calvin cycle broadly applied to almost all dicotyledons, but that it did not apply at all to plants as common as graminaceae, such as corn, sorghum and sugar cane, all of which are monocotyledons. These plants fix the CO2 of the air at a much higher rate. They need less organic fcrtilizer and they need less humus, because their carbonaceous components are derived out of the air in larger proportion. This is also the case for weeds.
In the Calvin cycle, the CO2 from the air attaches to ribulose 1-5 diphosphate molecules. Each resulting molecule decomposes into two 3-phosphoglyceric acid molecules. On the other hand, in the Hatch and Slack cycle, the CO2 is fixed by carboxylation of the phosphophenolpyruvate with formation of oxaloacetic acid. This acid reacts with C2 or C5 molecules to form pyruvic and phosphoglyceric acides.
This big difference between the two cycles can be presented in another way. In th Calvin cycle, the compounds formed from the CO2 in the air are primarily compounds with 3 atoms of carbon, such as glycerol-3-hosphate. In the Hatch and Slack cycle, C4 compounds are synthesized, such as oxaloacetate.
In other words, dicotyledons fix 3 carbon atoms and monocotyledons 4 carbon atoms. This explains the better yield for the latter. Synthesized enzymes are also different. There is more in the Hatch and Slack cycle than in the Calvin cycle. Some dicotyledons growing on salty lands follow the Hatch and Slack cycle. This would be due to a still unknown property of Na, tentatively attributed by some people to a transmutation of Na. These plants can be used to desalt a soil.
The Calvin cycle is the cycle in C3 and the Hatch and Slack cycle is the cycle in C4. Consequently, the latte produces more carbohydrates and more dry material. It requires only half as much water and more dry material. It requires only half as much water. This is the reason why these plants can grow well in fairly dry climates, as is the case with corn, sorghum, etc.
There is another important difference. In the C3 cycle, 40-50% of the atmospheric CO2 initially fixed is returned to the atmosphere by photorespiration. This could explain why in a closed chamber the atmosphere can still contain some CO2, although the incoming air has previously bubble through a solution of caustic soda to fix any incoming CO2. The remaining CO2 decreases slowly and asymptotically. The plant wilts and dies slowly. On the other hand, in the C4 cycle very little carbon is returned to the atmosphere. Almost all the carbon which is fixed is retained. Carbohydrates are formed more quickly. Grain plants, such as cereal plants, lack CO2 much sooner than most other monocotyledons when they are grown in a closed atmosphere, fed through a caustic acid solution. Their chlorophyll deficiency can be soon observed. At least, this is my interpretation.
(f) Isotopes ~
One should also observe in passing another phenomenon. Monocotyledons and dicotyledons are not only the seat of different chemical reactions, but they also have different physical behaviors. They fix stable carbon, oxygen and hydrogen isotopes differently when they come from CO2 and from water. In the sugar beet (a dicotyledon), saccharose does not have the same isotopic composition as the saccharose in sugar cane (a monocot). This question was studied recently with the greatest care. Its implication is not only scientific, but also economic. The selling price in sugar cane is much higher than the selling price in beet sugar. Fraud is possible. For the chemists in charge of the analyses, saccharose is saccharose; its formula is the same. It is impossible to determine its origin after it has been purified. But Nature does not dabble only in chemistry, it also dabbles in physics. C13/C12, O18/O16, H2/H1 ratio variations are high enough to be detected with mass spectrometry. Customs and the Agency for the Repression of Fraud are now equipped to check sugar to determine its area of origin. They use the same techniques with other compounds and other plants.
The treaties I know on photosynthesis are all silent on this point. Many other points are still unknown. Understanding of detailed reactions in chlorophyll still escape us. We only know some intermediary steps, and the molecular structures resulting from the combined action of photomorphogenesis and photosynthesis. We still do not know what happens inside the chlorophyll molecule at the atomic level. This molecule includes approximately 150 atoms grouped around a nucleus or heart of porphyrin. This porphyrin is formed around a magnesium atom linked to 4 nitrogen atoms. What happens at this level? Though magnesium is at chlorophylls core, it is never included in the formulas proposed by agronomists. Furthermore, it is assumed that chlorophyll preexists, but one does not known how Nature reproduces it. As the pant develops, the number of leaves increases, leaves grow and chlorophyll forms. Magnesium is a transition metal element, Does it act only as an electron carrier as iron in hemoglobin, copper in the hemocyanin of the shellfish, vanadium in ascidiae, etc.? There again we do not know the internal process generated by this central atom; we do not even know the process generated by porphyrin, which is built around this atom.
(g) Conclusions ~
Photosynthesis cannot be totally explained by chemical, hence molecular, phenomena, which have not been completely explored. There are indeed phenomena at the atomic level, such as variations in isotopic composition. However, we should consider the possibility of subatomic phenomena. These may be direct or indirect mechanism. I am inclined to say indirect and they work through the unquestionable and selective action of specific enzymes, growth hormones and other hormones. These compounds may contribute to the transmutations which were clearly shown in the course of the experiments. Nothing is simple. Results obtained with one plant cannot be extrapolated and applied to all plants. Failure will result if this point is not kept in mind. Each experiment should be performed according to a well defined procedure. In this field innovation may lead to poor results. Even a simple question of lighting, of inappropriate spectrum, may make the difference between positive and negative results. This is essentially the point I am trying to make.
(2) Photosynthesis (Limited to Traditional Aspects) ~
In most treatises the study of photosynthesis is based on the research based on spinach. This produces unwarranted generalizations.
In one kilogram of spinach leaves there are approximately 500 million chloroplasts. 60% of the weight of the proteins contained in the leaves are located in these small organs which are concentrated in the cytoplasm of the leaves.
These chloroplasts are 7-8 micrometer long and 2-3 micrometers thick. They are filled with a lumpy liquid, the stroma, which is surrounded by a double envelope. One of these constitutes a membrane network, the thylakoids. Thylakoids are vesicles filled with a liquid composed 80% of glycolipids and sulfolipids. Their envelopes contain very little phospholipids, but they contain chlorophylls (150 micrograms of proteins and 30 micrograms of pigments such as carotenoids). Nearly 50% of the proteins contained in the chloroplast are located in the thylakoid envelopes. Cytochromes and plastocyanin were found in them; they are proteins which can give or receive electrons according to their environoment. The nature of the constituents inside the thylakoids seems to be totally unknown as of 1980.
The stroma is rich in soluble proteins (about 0.4 gr/ml of stroma). It contains about 50% of the proteins in the chloroplast. The stroma contains the DNA and RNA which synthesize some proteins in the chloroplast. The lumpy aspect of the stroma, revealed by the electron microscope, is mainly attributed to ribosomes. The surface area for the external chloroplast envelope amounts to about 500 cm2 for one gram of leaf. The envelope contains only 1-2% of the proteins in the chloroplast. To this day, none of these proteins have been isolated it seems. Chloroplasts transform ADP in ATP under the effect of light and in the presence of electron acceptors.
The knowledge of photosynthesis progressed after intact chloroplasts were obtained in 1965. They simultaneously give some oxygen and fixed CO2 in a 1/1 ratio in the presence of light. However, chloroplast separation techniques were only applied to a very few vegetal species. Experiments were primarily performed with spinach. Test preparation was perfected for this plant. Its green material can be harvest all year round. Consequently, it was the subject of 70% of the research. This led many researchers to extrapolate their results to other species. This is a mistake. It was observed in some green plants, such as corn and sugar cane, that some intermediary steps were different from the steps in spinach reactions. This showed that the photosynthesis for spinach, a dicotyledon, did not apply to these monocotyledons.
When thylakoids are exposed are exposed to light, electrons are transferred between water and a final acceptor, of still unknown structure, designated as X protein. Walter Redox potential for the final acceptor, Eo = 0.1 V. Therefore, it is necessary to bring energy in, to operate the transfer. The energy is brought by light photons. This energy transfer is coupled with an ADP phosphorylation into ATP; this constitutes the energy storage mechanism. Light energy is collected inside the thylakoid membrane by two antennas each belonging to a different photosystem. These antennas comprise a system of proteins and pigments (chlorphylls and carotenoids) which absorb light energy and transmit it, by resonance, to the a-chlorophyll which is different in each photosystem. His chlorophyll is oxidized. One of its electrons is transferred to an acceptor. The transfer creates a high potential gradient, 0.8 V for one photosystem and 1 V for the other. For the first one, water is the electron donor, but to this day the acceptor is unknown. It is designated as the protein.
A magnesium base proteins intervenes during this reaction. Water is oxidized, giving protons and molecular oxygen (schematically: H2O + O = H2 + O2). The detailed mechanism of this reaction is still the subject of current studies. It is now generally accepted that oxygen is dissociated from water and not from CO2, as it was initially believed. In the other photosystems, the electron acceptor (X) is a protein which includes iron and sulfur atoms. Fe and S have a very low Redox potential. The two systems are connected by a chain of carriers which is little understood.
Q molecule electrons are successively transferred to quinines, to various cytochromns and then finally to plastocyanin, in which the electron donor is a blue protein containing copper.
Electrons are transferred from X proteins to ferroxoxin, which is a very small protein (molecular weight of the order of 12,000) of a low Redox potential. Eo = 0.32 C, containing Fe and S). Ferrodoxin contains iron and sulfur. Ferroxin can transfer its electrons to various molecules such as nicotinamide-adenine-dinucleotide-phosphate, NADP. It can also transfer them to a nitrite based flavoprotein. This transfer produces NH ions and electrons can be transferred to oxygen with production of superoxide radicals.
In this case the overall balance may be summarized by the following relation:
H2O + NADP >> NADPH2 + ½ O2
The NADP is reduced and the oxygen probably comes from the water. The primary effect of this transfer of electrons is the production of the highly reducing NADPH2 molecule; oxygen is only a by-product, so to speak.
We shall note that magnesium, the central atom of the chloroplast molecule, does not appear in these formulae. Its function has not yet been established. Many unknowns remain. Isnt such research purely speculative? Many teams of researchers throughout the world are attempting a direct transfer of electrons to a final acceptor by the channel of ferrodoxin coupled to hydrogenase. Hydrogenase is an enzyme extracted from bacteria or from molecular algae, through reactions producing molecular hydrogen. Hydrogen is a non-polluting source of energy. In nature, the ferrodoxin-hydrogenase pairing is observed among many bacteria produce a reaction similar to the above except that they dont free any oxygen. The thalla which live in symbiosis in the blue algae, does produce oxygen.
The production of hydrogen results in a pH modification on both sides of the thylakoid membrane. Plastoquinones, carriers of both protons and electrons, are reduced by Q molecules on the external face of the thylakoids and they capture protons from the stroma. However, the plastoquinones, oxidized by the cytochrome near the thylakoids internal envelope, also discharge some protons in the stroma. The protons formed during the oxidation of water contribute also to reinforce the pH gradient in the presence of light. Balance is due to the phosphorylation of ADP into ATP based on the formula proposed by the Englishman Mitchell:
ADP3- + H+ = HPO42- >> ATP4- + H2O
The detailed process of this schematic reaction is unknown.
ADP and NADPH2 molecules bring the necessary energy and the electrons to the stroma to fix the carbon dioxide from the air and to synthesize carbohydrates. 37 ATP molecules and 24 NADPH2 molecules are required for the synthesis of one saccharose molecule from H2O and CO2. In plant species of a type similar to spinach, the stroma inside the chloroplasts produces C3 molecules primarily (phosphorylated C5-oses: glyceraldehydes-3-phosphate or its isomer, dihydroxy-acetone-phosphate). This is a complex process realized in three steps. A C5 sugar is first decomposes in two 3-phosphateglyceride acid molecules. It is during this step that the CO2 from the air is integrated into organic molecules. One CO2 molecule and one H2O molecule are consumed. This reaction is catalyzed by an enzyme of large molecular weight (500,000). It is a complex enzyme composed of 8 large units and 3 small subunits. The second step is subdivided into two sub-steps and the third step corresponds to regeneration. This series of steps is known as the Calvin cycle or now, as the BBC for Benson, Bassham and Calvin.
Phosphorylated oses are produced in C6 (fructose), in C4, C7, and finally in C5 (ribulose). In 1973 Held showed that the pH of the stroma increased in the presence of light. The stroma loses protons to the inside space of the thyladoids until a balance is reached. In darkness it is the opposite. Enzyme activity is controlled by these pH variations.
Beet sugar and spinach leaves behave similarly. Both plants are dicots. Each square meter of beet leaves feeds about 130 mg/min of glucids in the veins of the leaves from which the glucids move to the roots for storage. The mechanism controlling the subsequent transformations of the 3-phosphates glyceraldehydes, after they reach the cytoplasm, is not known. Those which are not sent to the cytoplasm and those in the stroma, are transformed into starch which is a high saccharose polymer.
Contrary to saccharose, starch is not phosphorylated. There is a biological limit to the yield of the photosynthesis process. During the respiration phase, plants controlled by the BBC cycle waste a large part of the carbon integrated during the photosynthesis phase. It is estimated that these plants use only 1% of the 5.1020 kilocalories received each year from the sun. This consideration underlines the economic importance of the next cycle.
(3) Photosynthesis Cycle of Hatch and Slack ~
The photosynthesis cycle of Calvin is well known. It is the classic cycle in all published works, because it was most widely studied. In fact, a study of the C13/C12 isotopic ratio (research of 1968 by Bender) showed that it could vary by 5-15% and even more according to the photosynthesis cycle of the vegetal species. The study showed that this result was independent of the composition of the living medium for the culture and that it did not vary with time. The variations in the ratio could only be attributed to the physiology of the plant under the control of its specific enzymes. The emphasis was placed on the ratio between the two stable carbon isotopes in corn, as the chlorophyll metabolism seems to fix proportionally less C13 from the atmosphere.
Research on photosynthesis was first made with dicots because there was a convenient source of supply of raw material. The research extended to monocots such as corn. Sugar cane was included in order to stop fraud in sugar. Cane sugar contains more C13 than beet sugar. As previously mentioned, the Calvin cycle combines the CO2 from the air with ribose-1.5-diphosphate in beet sugar. The resulting molecule decomposes into two 3-phosphoglyceric acid molecules. As early as 1966, Hatch and Slack discovered another cycle in monocots. In that cycle, they showed that the CO2 from the air was fixed by carboxylation of the phosphenol-pyruvate and formed oxaloacetic acid. This acid combines with C2 or C5 molecules to form pyruvic and phosphoglyceric acids. In France, J. Bricout and others observed that the C13/C12 isotopic ratios were significantly different for beet and cane sugars. The decrease in C13 is of the order of 1.1%, compared to its concentration in the CO2 control, for cane sugar. The decrease is 2.5% for beet sugar.
We see the advantages of cultivating monocots rather than dicots. They derive more carbon dioxide from the atmosphere and require less organic feed. Many weeds have the same property and they grow even without fertilizer.
FIGURE 13 ~ [Missing]
FIGURE 14 ~ Values of Ca
Chapter 5The Devils Advocate
When I published my first works I had problems with chemists for whom theory precedes facts. For them, nothing is lost and nothing is created. This is true in chemistry. To many biochemists, life is only chemistry, and chemistry is the supreme science which explains everything, even the most abstract: will power, feelings, etc.
With a wealth of experiments I showed that chemical balances between elements in and elements and elements out were not always null. I was astounded by the naivete and the lack of realism of these biochemists. Their final argument was that a balance which was not null could not only be attributed to incomplete analyses. They claimed that some elements were surely hidden in a form which could not be detected by the analyses as performed. They did not realize that they only confessed their incapacity to perform correct chemical analyses. Chemists, even those who boast of being professors in our universities, could not be trusted.
Confronted with such common attributes, I just shrugged my shoulders. To avoid useless polemics, I decided to stop referring to chemical analyses and to use modern techniques of physical analysis such as flame spectroscopy for J, atomic absorption spectroscopy for Ca, etc. At the beginning of my research, physical techniques to quantitatively measure specific anions, such as S, P and N, had not been developed. The same is still true 20 years late. His is why my studies, and those of researchers who wanted to duplicate them, were limited to the study of balances for a few important cations by various physical techniques.
In France a university professor, who was a high-ranking Rationalist wrote in a publication that I was wrong. He was director of the biological chemistry laboratory in his university. Rationalism was a philosophy publicized by Robespierre, pope of the Goddess Reason. This professor had decided to duplicate, as he said, one of my experiments. In fact his experiment differed extensively from mine. A summary of his study was included on pages 88-91 of my book, now out of print, Preuves Relatives a lExistence de Transmutations Biologiques, published by Maloine in 1968. The essential argument was reproduced on pages 212-213 in my book of 1975, Preuves en Biologie de Transmutations de Faible Energie, which is still being reprinted. I will not say more on the obvious bad faith and the exaggerated cynicism of some sectarian objectors. Far be it from me to place all Rationalists under one label. There are exceptions among them as there are exceptions everywhere. I learned that several distinguished scientists among them protested against the attitude of this learned master.
However, he was not the only one to think in this way. I mentioned the case of the hidden iodine produced by some algae, which cannot be measured in a certain vegetal state, but can be measured in another state. This showed indeed that some experiments were deficient. To point at the errors was not sufficient. To disdainfully reject gratuitous affirmations was not the solution to definitively eradicate affirmations was not the solution to credulous and unscientific minds of too many biochemists. Figures were necessary. In the book mentioned before, dating from 1968, on p. 158-167, I had indeed given some precise information. However, it concerned phosphorus variations during the germination of lentils. Undoubtedly this escaped various phytobiologists. I noted in a publication, Revue de Biologie, that a professor of botany was still imbued with 19th century dogma in 1980. Criticizing the data presented by Zundel on the increase in Ca during the germination of oats, she writes, "If the ashes are dissolved in hot HCl, the acid cannot dissolve Ca which is in the form of sulfates and phosphates. An underestimation of the total quantity of Ca in the caryopses could result, whereas the Ca contained in the seedlings, and which can be oxidized, would be counted in its totality. This could explain the increase in Ca which was noted". General statements are made without regard for the precise experimental data, which are not even mentioned. In the previous paragraph, she wrote: "In caryopses, Ca is partially found in phytins (inositol hexaphosphate of Ca and Mg) in aleurone granules, and partially as insoluble phosphate and sulfate. In seedlings, Ca is found mainly as pectate, which is transformed in oxide by combustion".
But what are these pectates? "Combustions"? Arguments, but no figures! The "partially" is meaningless. Which part? Which proportion? I will quote figures later. So much for the questionable, and even partially wrong. Opinion expressed by this university professor. Before presenting my figures, I believe I should say a few words presenting my figures, I believe I should say a few words about phosphorus which is not part of this work.
Chapter 6Problems Related To Phosphorus, Complexity Of Phytoanalyses
Phosphorus and calcium in living matter are closely related. They have been studied for a long time. I dedicated a chapter of my 1975 book to Vauquelins studies on the balance of the calcium ingested by a hen. Calcium is found as carbohydrates and sulfates in oats, in the egg laid, and in droppings. Vauquelin mentioned chemical analysis procedures to measure the Ca linked to P and to the carbonate. This is not a new problem.
In my book of 1968 I presented some detailed information on the studies related to form changes of P during seed germination. It came from a doctoral thesis written in 1935 by a pharmacist, Yves Colin, after a five-year study. I own a printed copy of this document in which 84 pages are dedicated to "Technical Research on the Separation and the measurement of Primary Phosphorus Elements in Seeds". Colin presented some data on wheat and on sunflowers, but his research was primarily on lentils. P and Ca contents are the same in lentils and in oats. However, there are marked differences in the nature of P compounds in different vegetal species, so that the analytical procedure varies from species to species. I did not include in my 1975 book the chapter which dealt with this matter in my book of 1978 which is now out of print. One can refer to Y. Colins thesis in university libraries. It will show precisely how wrong and biased were the statements expressed by the professor of botany mentioned previously, statements unsupported by quantified data.
Good chemists know that P compounds are not soluble in any old solvent. Solvents must be selected. Care must be brought to the sequence of the reactions. A reaction can stop the sequence by fixing P in a compound which is then totally insoluble in the following reaction and it will then resist all attempts to measure it.
In plants, P is found in phosphoric ester such as true phosphoro-lipids. Among the photo-lipids are found lecithins, but mostly they are glycerol-phosphoric acid derivatives. One also finds P in phosphoric acid esters of sugars, diphosphoric hexose monophosphoric ester, momophosphoric polyoses, etc. During the last decade of the 19th century and the beginning of the 20th century many studies dealt with P compounds and their quantitative variations in germinating pants. Phospo-lipids and phospho-amino-lipids were studied first, then the nucleic acids which incorporate large quantities of organic P were studied. After hydrolysis, they release o-phosphoric acid. Nucleic acids differ according to the plant; some information on this subject may be found in my book of 1962. These studies led researchers to look for various ways to separate these compounds. For a given solvent, results may vary according to acid concentration and temperature. My former colleague at the Conseil dHygience in Paris, Gabriel Bertrand, was convinced that transmutations occurred in the metabolism of living matter. He also showed me that P contents in ashes and in the dry material were different. Differences were of the order of 10% if reactions, on material dried at 100° C, were performed in a wet medium. At 200 C a loss of P was observed: this was mainly due to the fusion of of alkaline phosphates. Fusion produced a slag surrounding the particles of organic phosphorus which could not be decomposed by acids. Generally, these phospo-lipids and phospho-nucleic compounds have no close relation with Ca; their study may be ignored in research on Ca.
On the other hand, phosphoro-proteids, which are not water-soluble, but are alkali soluble, are very important in fresh plants, though they are not widely found in animal tissues. Phytin was discovered during the 19th century. It is a hexaphosphoric ester of inositol, or more exactly it is a Ca and Mg salt of this ester. In 1903, Posternak established that organic P accounted for 22% of the phytin, Ca for 12% and Mg for 1.5%. therefore there are large quantities of Ca. Inositol is a polyaclohol widely distributed throughout plants. In grain plants it accounts for approximately 50% of the total P and over 70% in sunflowers. In his thesis, Colin describes the techniques which were perfected since the beginning of this century. These techniques were primarily developed by Javillier around 1930; we both sat together on the Conseil dHygience. He modified the analytical technique developed by Copeau in 1927. Mineralization comes about through a mixture of sulfuric and nitric acids (to which Baranger added some concentrated hydrogen peroxide). Phospho-amino-lipids are first extracted. Some per-compounds are explosive and must be handled with care. One must follow carefully the instructions given by the specialists in this type of research. For our study of Ca, the important analyses concern acid-soluble P and then mineral P. To separate phytinic P from total P, Colin had to develop a technique derived from Javilliers procedure. This was because the P in the seeds of cereal plants differ markedly from the nucleic acid formed in lentils. The presence of iron modifies the sequence of the reactions and requires the use of hydrochloric acid.
In 100 grams of dry powder of lentil seeds or of oats, there are 300-400 mg of P in total. 50% of this total is in phytin; a little over 10% is lipid P and a similar amount is nucleic P. The ratio of the last two kinds varies little during germination. As they are not linked to the production of ca, we will not show any data for them. Colin quotes results for batches of 400 lentils picked every third day after the lentils were prepared for germination. Here are the weights in mg for each batch:
P (Phytin) ~ P (Mineral) ~ P (Total)
Before Germination: 50.63 ~ 18.30 ~ 92.00
After 3 days: 38.31 ~ 30.84 ~ 93.65
After 6 days: 12.64 ~ 54.61 ~ 90.00
After 9 days: 0 ~ 62.30 ~ 87Other analyses were made after 12 days and after 33 days. We will not discuss the results here. The main point is that the P in phytin decreased significantly after 3 days and that it decreased by approximately ¾ after 6 days. It is around the 4th and 5th days that the rapid transformation of phytin P into mineral P takes off. There is also a progressive decrease in total P, which was confirmed by other studies on soybeans and on vetch and oats. I shall come back to this point. It is remarkable that all the organic P in the phytin disappeared after 9 days, as it was transformed into mineral P.
In the seed, mineral phosphates of Ca and Mg amount to about one-half of the P contained in phytin. The Mg is negligible. In the plant there is no more phytin after 9 days and mineral P is a little over 3 times greater than in the seed. Therefore, if the mineral phosphate in the plant is neglected by the experimenter because it is not soluble in HCl, the amount the amount of Ca in the plant is underestimated. This is the opposite of what was stated by the professor of botany mentioned in the previous chapter.
Chapter 7How To Correctly Duplicate A Typical Experiment On Biological Transmutations
When one wants to check a phenomenon, witnessed by another person, it is not right to give an opinion on the validity of the phenomenon unless the procedure used to establish it is strictly followed. If one parameter is different and if the authors conclusions are not confirmed, it is right to first ask of the failure of the experiment is not due to the change in the procedure and to the neglect of some essential points.
In other words, the verification of an experiment requires the exact duplication of the experimental conditions set by the researcher who established the existence of the phenomenon. It is legitimate to innovate only after this has been done.
Too often operators attempting to duplicate experiments modify the procedure described by the inventor. They do so for several reasons.
In the goal they pursue they underestimate the importance of some points of an experiment, which is new to them. This may be due to ignorance or to professional bias. Some extremely complex fields of research require a good knowledge of nuclear physics and chemistry, and an outstanding knowledge of biology. This is the case for biological transmutations which are biophysical phenomena which can be detected by wet chemical analysis or physical chemical analysis.
(1) Study of the Calcium Variation in Cultivated Oats ~
We will look at a study performed by a team, composed of a physicist and a phytobiologist, in an American university.
They wanted to check if there was an effective increase in Ca and an effective decrease in K during germination as compared to the initial amounts in the seeds. Germination took place in ultra-pure water. No Ca could be derived from the water nor from the bottles used for the experiment. This was checked prior to the beginning of the experiment.
Theoretically, according to classic rules, the Ca balance, as well as the K balance, should not change, because "nothing is lost, nothing is created". This is true in chemistry. However, this does not apply to some specific biological processes involving not only chemical reactions but also prior to them, phenomena ruled by nuclear physics. Chemistry deals with phenomena involving electrons, and in general only the outer shell electrons of the atom. Electromagnetic energies, it would seem, cannot cause any transmutations. There are various reasons for this, but they will not be discussed here. These transmutations can happen under the effect of strong or weak interactions as it will be explained in Part II of this book.
After performing only one experiment, these American scientists concluded that my observations were in error. For the K/Ca ratio, they found an average value of 4.5 for the seeds and only 1.5 for the plants after germination and after too short a cultivation period, in my judgment. From the outset, they should have attributed K/Ca variations either to a decrease in K, or to an increase in Ca, or to simultaneous variations of K and Ca. Lets more closely examine the reasons which led to their conclusions.
They analyzed three aliquot parts of a batch of seeds and of plants grown from "identical" seeds. I will not cast any judgment on this technique, which, it seems, was used for the first time in this case. It might be a mistake to try a new technique in this instance. Though I know of so many scientists who are prisoners of their habits and sometimes of their equipment and who are incapable of admitting that they are wrong. To them, their technique is the best; they adopted it once and for all. This is a conceited attitude common to too many scientists. It is a mistake. Various techniques should be used for cross-checking.
For example, I observed some unacceptable discrepancies in analytical data obtained by complexometry techniques for Ca. The nature of the complexing agent must be adapted to the Mg/Ca ratio in the solution analyzed. In spite of a double precipitation, the same is true for analyses by means of oxalate salts. Specific techniques are legitimate in some cases and not in others. In the same way, precipitations must be performed in the right sequence. Otherwise a particular molecule may be made insoluble to the detriment of subsequent reactions. Unfortunately knowing and remembering this point is not enough. This is the case in experiments involving biological transmutations, when the outcome of a first transmutation is unknown in advance. A technique may be valid for the control and not for the end product due to the modifications caused by the transmutation in the ratios between some elements. Two steps of an experiment may not always be cross-checked with the same technique if one wants to reach the right conclusions.
Of course I do not object to chemical analyses, but the point emphasized above should always be kept in mind. I always used the atomic absorption spectrometry technique for the analysis of Ca (or Mg, Fe, etc.) to obviate the uncertainties presented by some chemical techniques and to prevent (often ungenerous) objections by experimenters imbued by their own technique. Uncertainties in chemical procedure can generally be cleared, once the experiment is mastered and once its results have become reproducible. This requires that the parameters of the experiment be maintained constant. Zundel used a chemical technique, developed by Prof Charlot, for the analysis of Ca in oat seeds and plants. The material is heated to 950° C to avoid the formation of carbonates, and to insure that all Ca in the final reaction is in the form of CaO. The CaO is weighed to 1/10 mg for each batch. The amount of Ca is then computed. With this technique, Zundel obtained fairly constant figures, though they were always slightly higher than those given by atomic absorption spectrophotometry. This was true for both seeds and plant so that the ratios between the Ca contents in the seed and in the plant are independent of the analytical techniques. Variations in the absolute value of Ca may be ascribed to various causes: calibration differences in the physical technique or the presence of contaminants not precipitated previously in the chemical analysis. Despite these factors, comparative results obtained by these two techniques often show little difference. Balances are usually reproducible to a few percentage points with each technique. Too many physicists, even chemists and biologists tend to ignore, underestimate or neglect differences from one biological batch to the next. Among seed as among animals, there are vigorous subjects and there are weaker subjects. At times differences in the nature of the subject can be detected chemically or by analysis of the isotopes. The isotopic composition of an element does not necessarily translate to an organic compound of the same element. The chemical formula for its three main components, H, O and C, are not the same for cane sugar as for beet sugar. In biology calculations should not be based on standard atomic masses, which are average values as stated in our books. Although they are precisely defined, they are only arbitrarily selected standard values. In reality there is no such thing as isotopic selectivity.
This is why, in my studies, I decided to accept only variations greater than 5% in average values for the elements considered. It is understood that the number of analyses performed must be sufficient to yield statistically valid averages.
Several cross-checks by neutron activation performed in authorized nuclear physics laboratories showed that the technique was also valid for Ca. Magnitudes were approximately the same as those found by atomic absorption spectrophotometry. With a minimum of 5 readings, measurement dispersion was smaller than 4%. Neutron activation, which is non-destructive, should not be used to first test a seed and then the palnt grown from this seed. I was able to establish that the plants germinating factors were altered by this technique. The plant metabolism is modified and transmutation, which are produced by enzymes, are slowed down. (please refer to my book of 1975, p. 233).
From the beginning of my research 20 years ago, I abandoned chemical techniques for K analysis. Prof Baranger of Ecole Polytechnique in Paris, a chemical analysis specialist, finally abandoned the chemical analysis of K after he had obtained questionable results for several years. Zundel, a chemical engineering graduate from the Zurich Polytechnicum, who supervised a chemical analysis laboratory, also abandoned the chemical analysis of K from the start of his studies on transmutation during the germination of oats. We used emission flame spectrometry despite the inherent difficulties in reducing measurement discrepancies from one sample to the next.
The flame spectrometer is still an instrument which is difficult to set for good reproducibility. It is now used by everybody, as there is no better instrument for the various purposes. Many readings are required with this instrument to get statistically valid results. There are some reservations on the various statistical methods prevailing in different countries as all these methods are based on arbitrary postulates.
It is impossible to obtain statistically valid conclusions on biological samples from a small number of analyses performed on aliquot parts, which are sometimes too light in weight to start with. Usually we make 5 to 10 readings on a given sample. At times we made up to 50 tests. Baranger tested up to 300 and even 400 samples one by one. On the other hand, it seems that only three analyses were performed in the USA. The results obtained were so scattered that they were statistically invalid. Nevertheless, it was the basis for the negative conclusions expressed by the scientists who performed the experiment even though the average data showed a large variation of the K/Ca ratio.
(2) Biological Conditions ~
Ca and K variations during the germination of oats observed in the USA were different in absolute value from those we had customarily observed during tens of congruent experiments. These experiments included hundreds of analyses performed on thousands of plants and tens of thousands of seeds. This consideration led us to carefully examine the content of the report written by these scientists.
It showed us also that they underestimated or ignored some biological factors.
We recommended oats for the study of Ca, as this plant was extensively studied and it showed clear results. The increase in Ca is considerable, often greater than 100%, independently of the batch or the species, when the seed is germinated in twice-distilled water with an acid pH (5.6-5.8). The oat plant grows on acid soils, hence it is clearly a calcifuge. The seed contains little Ca, an element necessary for the first reactions for the germination process. Ca is required until growth hormones are synthesized. After 8-10 days the total Ca content of the seed and the seedling begins to increase very rapidly at the expense of K, although no Ca can come from the outside. The seed is rich in K as its K/Ca ratio is approximately 4.5 and varies little for different species. It constitutes the reserve for the growth of the plant. This ratio decreases the reserve for the growth of the plant. This ratio decreases to about 1.8 over 3-4 weeks and then declines asymptotically to 1.5, the minimum value reached after 6 weeks. After this time the K reserves are too low to compensate for Ca and the pant starts to wilt. We do not know if the pH was measured in the American study. The pH is not mentioned in the text we received. It is not critical in this particular case, because the water pH was certainly acid.
It seems to me that unsatisfactory cultivation conditions are the primary reason why no statistically valid increase in Ca was found. A researcher should not try to innovate while exploring entirely new grounds. He should follow the instructions of the people who have long experience with the phenomenon and who have mastered this phenomenon only after extensive research.
My attention was drawn to a short sentence mentioning that the germination was performed under artificial fluorescent lighting with a lamp of the warm white type. As I have studied fluorescent lighting over 20 years, I jumped. The warm white lamp gives a spectrum which appears close to the solar spectrum to our eye. It enables agronomists to easily detect color changes in the seedlings. However, such lamps should not be used for photosynthesis. Plants and the human eye have very different sensitivities to light spectra.
In the beginning our studies were only made under sunlight, to approximate natural conditions for the plant as closely as possible and to prevent the introduction of an additional unknown. After a few years, various considerations forced us to use artificial lighting. We needed to experiment at various times and in various seasons at poorly lit locations. Some varieties of oats, often called winter oats, are sown in fall, others at the beginning of spring. It took years of research and the help of discharge lamp specialists to select the right lamps. Only these lamps could radiate the short wavelengths in the blue, violet and UV which are so important for photosynthesis. Wavelengths in the red, which carry heat, are also well represented in the spectrum of these lamps. On the other hand these lamps radiate very little in the yellow, the band of maximum sensitivity for our eye.
This research led to the development of lamps with spectra well tailored to photosynthesis. For visual observation, lamps of the warm white type or equivalent are needed. However, they are only used intermittently and they cannot replace solar light in any way for photosynthesis.
The phytolamps used were usually rated at 250 watts and sometimes at 400 watts. This was a high but necessary power rating. These lamps radiate a great deal of heat and they must be located outside the cultivation chambers. The chambers are not made of glass, but of metacrylate. For supply considerations Altuglas rather than Plexiglass was used. Metacrylate, as glass, absorbs some UV wavelengths. The bands filtered change according to the material. Therefore, plants cultivate under glass or metacrylate are affected differently by light photons than plants grown in the open under sunlight. For this reason we placed a small lamp inside the chamber rated at a few watts, which radiated complementary wavelengths in the near UV. With this lighting, we obtained plants as vigorous and as rich in Ca as plants grown under direct, unfiltered sunlight.
The American researchers stated that their seedlings were more yellow (chlorosis) and more flaccid than those normally grown in sunlight. They did not mention if they had considered the possibility of deficiencies in photosynthesis. Another effect of this process is easily checked. Under this mechanism, the plant produces some dry material from various synthesized carbohydrate compounds (glucids, lipids, parotids, etc.). Lignin, composed of the resulting celluloses (hemicellulose, etc.) contributes to plant system rigidity. The stems of the plants grown by the Americans remained flaccid and bent. This was a sure sign of deficient photosynthesis. In order to form carbohydrates, the plant takes its carbon from the carbon dioxide in the air and its H and O from the water absorbed by its roots. A satisfactory photosynthesis is therefore indispensable to plant metabolism and for the synthesis of essential growth enzymes; it is absolutely necessary for the good health of the plant. An enzyme linked to oxidizing phosphorylation, and created at the internal surface of the mitochondria inside the envelope, is responsible for the formation of Ca from K with Mg as a catalyst. The enzyme active in this reaction is derived from ATP chelated with Mg (Mg-ATPase). This is a point I discussed in my books. Another point was presented by S. Goldfein in a report written for and published by the scientific section of the US Amy in Fort Belvoir, CA in 1978. Without the normal activity of this enzyme, there is a deficient plant metabolism and a transmutation which is too weak to yield significant amounts of Ca. This happened in the experiment performed in the USA. The seedlings were weak and stunted. The stems were only a few centimeters long. During the same growing period, we obtained firm and erect stems 20-20 cm long. We also obtained a much greater weight for the dry material, and even for the ashes, than for the seeds. A poor photosynthetic action prevents the formation of carbohydrates. Furthermore, the weights for the dry material or the ashes are smaller than the seed weights because of the exhaustion of the seed reserves due to the release of CO2 in the atmosphere during respiration.
Another point deserves attention. In a controlled atmosphere, the incoming air should contain some CO2 as a source for the photosynthetic process. To prevent any external Ca contamination, this air must be filtered. It should be bubbled through an HCl solution contained in a flask, so that any trace of Ca, which escaped from the filter, would be retained. To neutralize any trace of acid carried by air bubbles, it would then be bubbled through a flask containing a basic solution. The base should not be NaOH which would absorb CO2. NaHCO3 was used because it lets CO2 go through freely. We noted no clues in the report we received from the USA which would lead us to believe that these precautions were ever taken. After scrubbing in acid and basic solutions, the incoming air is scrubbed in a flask containing pure, twice-distilled water before it lets CO2 go through freely. We noted no clues in the report we received from the USA which would lead us to believe that these precautions were never taken. After scrubbing in acid and basic solutions, the incoming air is scrubbed in a flask containing pure, twice distilled water before entering the closed chamber. The chamber is maintained at a low positive pressure of about 3 mm W.G. to prevent any unwanted air intake through the gasket.
We will mention another study related to the function of CO2. Photosynthesis develops very poorly without this gas. It is only active for a very short time in the morning when the plant absorbs the CO2 it released during the night by respiration. During the night the plant draws on its reserves of oxygen and carbon and in doing so, becomes weaker. The period of activity corresponds to the time the light is turned on each day when artifical light is used. The inverse experiment was also performed by increasing the CO2 content of the air supply to the chamber. Zundel showed that a large increase in the CO2 concnetration (about 10 times the normal concentration in atmospheric air) led to an unsatisfactory plant metabolism. There is excess carbon. This requires an abnormal increase in both the absorption of carbon and in the decomposition, which generates hydrogen and oxygen. As vein sections are too small, the plant weakens and grows sickly. On the other hand an increase of 2-3 times the natural CO2 concentration in the air is beneficial. Some algae, chlorella for example, are cultivated in atmosphere rich in CO2.
At this range of CO2 concentration we observed a new phenomenon. The increase in Ca was very large (500-600%) and it could not be ascribed solely to the decrease in K. Analyses showed that Si decreased very significantly, but insufficiently to account for the increase in Ca.
This phenomenon does not occur in a natural atmosphere as was shown in the work with oat plants. Various analyses, chemical as well as physical, supported this. In atmospheres overloaded with CO2, there was a large decrease in SiO2 stored in the seed. This was not entirely surprising. We knew that in Nature there was a partial inverse compensation between Ca and Si under specific conditions, which have not yet been completely studied. We studied the reaction Si14 + C6 >> Ca20 in various books.
This explains why horsetail (equisitum) silica was used in phytotherapy in antiquity to recalcify patients. It was necessary to harvest the horsetail at the end of spring, therefore at the end of the period of full growth, so the silicon would be present as part of organic compounds and not under the prevailing form of SiO2. Silicon is stored as mineral SiO2 in the rhyzome of the pant during fall and winter. Similarly, I made a detailed presentation (See Preuves en Biologie...) of the research by the great French chemist Vauquelin on the balance of Ca in laying hens. These hens were only fed oats. "Lime" was measured after precipitation of the carbonates and phosphates contained in the oats and in the excretions from the hens (eggs and droppings). A large increase in the "lime" balance was noted during these experiments. Vauquelin thought that this "earth" or lime (we would call it today an alkaline mineral compound) could only come from another "earth" contained in oats, silica. At the time, knowledge of the atom was nonexistent. He could not imagine that the K19 + H1 >> Ca20 reaction was possible. During his measurements, he observed that the increase in CaO was greater than the clearly measurable decrease in SiO2. From this he concluded with a very modern rational that Ca could not come totally from Si. The problem still has not been resolved. We see how long ago (1799) the inverse relation between Si and Ca was noted. We will not discuss here the hypothesis which would explain the presence of Ca as coming from the excess CO2. Research on this subject is still too speculative.
(3) Conclusions ~
We showed some of the weaknesses in the studies made in the USA, or at least in the documents which were forwarded to us. Even trained researchers with a background in a different field can make mistakes. For example, Prof Baranger decided to limit his study to a single vetch species, Vicia Sativa. He selected it for a reason of convenience because the seeds were very nearly round and easy to gauge, without any regard for the vegetal biology aspect. To achieve great accuracy in his measurements, he would weigh 10 seeds from a Petri dish to 1/100 mg, which is difficult. Each batch of control seeds was weighed separately. So was each batch of plants, growing on two layers of extra-pure and ash-free paper, moistened with twice distilled water. For the integrity of the experiment, the data obtained on the batches of seeds and on the batches of 3-4 week old plants were handed over to a mathematician, specialized in statistics. Baranger always analyzed a minimum of 100 batches of seed controls and of 300-400 batches of plants. The data obtained from these hundreds of analyses showed that in average, there was a significant increase in Ca in the seedlings during germination.
However, this increase in Ca, which was statistically established, looked too small to me. It was neither impressive nor spectacular. The increase was usually in the 2-3% range, and seldom did it reach 6-9%. I always express reservations on the 1% accuracy level in the chemical analysis of biological materials. Batches of seeds age. Batches extracted from a given stock over a period of one year are not endowed with the same vitality during their respective experiments. Even a germination rate of 95% or more does not guarantee 1% accuracy. Seeds, as with individuals of a same species, have different innate strengths; their enzymes are not equally active. Averaging the data from hundreds of experiments obviously alleviates the problem created by these differences. After selecting vetch seeds for handling convenience, Baranger looked for optimal germination conditions. He noted that in twice distilled water the increase in Ca was very small. It increased somewhat up to 6-9% when a Ca salt was added to the water. After trying various salts, he selected calcium chloride (CaCl2). The additional Ca was added to the Ca content of the seed. The sum was then subtracted from the sum of the Ca contents of the plants and of the filter paper impregnated with the excretions from the plants. Prof Baranger had to recognize that seeds, stored for a long time, became moldy as soon as they were dampened. Should then the transformations be ascribed to the molds or to enzymes in the seed?
We noted that Baranger never mentioned the pH of the culture medium in is reports. He found that additional Ca strengthened the plant, but he apparently was not aware that this effect was a calcicole property of vetch, which grows better in the presence of a little Ca, in an alkaline soil. This is not a clearly calcifuge plant, like oats, which requires an acid medium, a generator of protons, in order to produce the reaction. Vetch does not have the enzymes required to form the additional Ca needed for plant growth. There ism hence, a very small increase in Ca in the plant, usually less than 5% in average. On the other hand, on the oat plant, the increase is sometimes greater than 100% when it is cultivated in an acid medium. Such is not the case when the variations observed are 5% or less. For example, Barangers research on P seems to show a decrease in P in the range of 1-1.5%. This is questionable even if the precautions taken tend to validate these figures. I should add that Barangers report does not give any indication that he was worried about the artificial light spectrum for the glass case he used in his experiments. When I visited his laboratory, he had no phytolamps with spectra adapted to photosynthesis. He used a Daylight-type lamp, adapted to human vision. The few experiments performed in sunlight were made under a glass cover which protected the medium from falling dust, but not from floating dust. His seedling trays were located in a narrow courtyard, or rather a ventilation well, next to his laboratory. There was no direct sunlight at all. All his seedlings were deficient in chlorophyll. This might explain the small size of the variations obtained.
In spite of every precaution he can think of, a specialist in chemistry and biological chemistry may overlook some aspects of an experiment, because he is too specialized. Yet Baranger established, in the course of 10 years of research, that in Nature there were variations in some elements which could not be explained by the theories included in official teachning programs. Nature can do more than chemistry. Baranger did not want to hypothesize on the causes of the variations. He did not want to go against established nuclear physical theories and he chose to limit himself to his specialty and to remain exclusively in the domain of experimentation. However, we see that it is necessary to leave traditional practices and to use a large number of different techniques which are often difficult to unify.
It was even more difficult for two specialized researchers in the USA to reach valid conclusions after making only one series of experiments on small batches (usually 30 seeds each) and to avoid major pitfalls.
Final Remarks ~
I would like this last experiment to be duplicated again in this university or another. Then the points made in the present document could be taken into consideration. As preparation, I believe that it is indispensable to perform a series of experiments on oats only, as this plant was the subject of many studies. These studies allow the experimenter to locate errors, as there is by now a solid database for oats established over a long period of experimentation.
For health and age reasons, I cannot travel any more and I declined invitations to travel in the USA, from the Los Angeles Park Service, for example. Following experiments based on my work, the Park Service modified its application of fertilizers. The concepts of classical agronomy have been borrowed directly from chemistry. They lead to excesses or deficiencies injurious to the plant. What the plant needs is not what is found in the mature plant, at the end of its metabolism. We saw that this was true of Ca. It is also true for other elements. These needs vary according to the soil and the plant species (calcicole or calcifuge, for example). The whole of agronomy must be reconsidered. The same is true for animal husbandry, and human and animal dietetics. Many dietetic or medical publications deal with applications of transmutation, a property of living matter. These transmutations are produced under conditions controlled by nuclear physics and unrelated to the strong interactions which lead to the atomic bomb. The Life process is gentle. Those of my works which were published after1974 show how this phenomenon can be integrated in the physics of neutral currents or weak interactions.
Note: Other experiments were duplicated in the USA in a school, in which a student presented a paper several tens of pages long each year. The studies were related to several kinds of plants (oats, wheat, soybeans, barley, gumbo-okra) and to various elements (P, Mg, Ca, K, Na). I did not mention them before, because it seemed to me that the students overlooked some important parameters. They did not contact me, nor did their professors. I only received the data later. I did not have sufficient details to judge if the excessive dispersions of the results should be ascribed to errors in the growing procedure, to mistakes in the preparation of samples to be analyzed, or to any other causes.
The same is true for the studies directed by Komaki. I cannot accept the figures I received for various elements in different kinds of pants: rice, soybeans, garlic and azuki, a species of small beans. In fact, all these studies show that the respective compositions of a plant and of its seed are always different. Also in France working groups of students preparing for the BTS exam (Brevet de Technicien Superieur) in agronomy proceeded to do various studies. However, the most important precautions mentioned above were not taken. I could not comment on the variations observed in the data. The same was true for studies made in Switzerland, Germany and other countries. This is why I thought it important to publish the information presented in this document.
Figure 13 [Not included here] ~ Groupe de travail dune Ecole Technique Superiuer a Nantes, en 1976, effechant des researches sur la variation du Ca dans une avoine de variete Peniarth
Chapter 8
Mass Spectrometer Analysis
In the second half of 1979 I learned that the microanalysis laboratory of CNRS just acquired a mass spectrometer of a new model linked to a computer. It could measure the elements quantitatively, as previous instruments gave only rough quantitative indications: "traces", or if the line was clear, the designation of the element was followed by one, two, or three stars. Therefore, I had never been able to cross-check positive ion contents by any spectroscopic method. The determination of the P content was important as it was accepted that the P/Ca ratio was close to 606.5 for oats when P was determined by chemical techniques. Although I have described above some aspects of the complex quest for various P compounds, I ignored some other points. For example, the presence of manganese prevents the analysis of P. It is eliminated before molybdate is added. The Mn content in oats is small and it is close to the Fe content. A part of the P, remaining in the form of undetermined compounds, is ignored, but it has little effect on the total results of the analysis.
For better accuracy, it was necessary to eliminate the error on the low side which was due to seed and plant incineration. For this reason, I requested from CNRS a mass spectroanalysis of various positive and negative ions, in order to cross-check the results obtained by the other techniques. We sent to CNRS powder samples obtained after drying the material at 85° C for 72 hours and grinding it. This was below the limit of 100° C recommended by Gabriel Bertrand, the temperature at which he operated. By working at lower temperatures, the vaporization of some organic P compounds and the formation of alkaline slags, insoluble in subsequent reactions, were prevented. The data presented below for P and Ca only are given for cross-checking purposes. Lets keep in mind that only one cross-check experiment was made, and that the values represent the average of two measurements. I received a written confirmation of this data in early January 1980. The figures are expressed in milligrams of the particular element per 100 gr of dry material.
S/P/V: P ~ Ca
Seed: 485 ~ 76
Plants: 310 ~ 115.5
Variation: =175 ~ +39.5The P/Ca ratio is of 6.5, therefore normal. Consequently, I believe the calibration to be correct. This analysis, which was performed in a French laboratory, does not represent an absolute proof. The experiment should be repeated many times. As I was only looking for a cross-check and, as the data was compatible with the data obtained by other techniques, we may accept the data as reliable. They confirm once more that some variations occur in the amounts of some elements during the germination of oats, although no additional amount of these elements was brought from the outside during that period. It was not the purpose of the CNRS experiment to prove that the increase in Ca was linked to the decrease in P. Experiments of a completely different type would be needed for that. It is obvious that there is no quantitative relationship between the absolute values for Ca and P. These discrepancies cannot be attributed to errors in the experimental procedure as there are increases for some elements and decreases for others. The figures tabulated above are not absolutely accurate, because the dry weight of the seed, from which the seedling originates, is not exactly the same as the dry weight of the control seed. The average weights for seed and seedling were computed over hundreds of trial. These computations showed little quantitative variation and no qualitative variation in the data shown above. This is the reason why I do not reproduce here the details of these experiments. They were published in the scientific press in French, English, and other languages.
As the quantitative bases are different, it is not legitimate to compare percentage variations, as some people are tempted to do. After standardization, there is a 32% decrease in P and a 52% increase in Ca. It would be absurd to state that there is an appropriate compensation around 44 ± 8%.
After receiving a confirmation of the P concentrations in oats at the beginning of 1980, I decided to further investigate the relative importance of the Ca which is insoluble in HCl. This is Ca linked to the phosphates. I wanted to use these new figures to prove wrong those who still insisted that the variations in chemical balances came from inappropriate chemical analysis procedures. These statements were unwarranted: furthermore, they contradict the conclusions drawn by their authors. Zundel performed three additional experiments on a total number of 870 seeds and seedlings. He gave me the results in late November-early December 1980. I received enough data to be able to form an opinion. Peniarth oats used were supplied by INRA. The seeds weighed an average of 34.21 mg each. Seeds and seedlings were then incinerated at 950° C and chemical analyses were performed using Prof Charlots technique. The weight of the material which was insoluble in HCl was 0.013 mg. The amount of Ca in this material was then determined. The following results were obtained:
Ca content in insoluble material per seed: 0.0032
Average Ca content in insoluble material per seedling: 0.0319The second figure is equal to almost 10 times the first one. This confirms that an important molecular transformation happened, and it transferred a very large quantity of Ca into the insoluble state. Neglecting the Ca linked to the insoluble material leads to a very sizable underestimation of the Ca production during the germination of oats. This contradicts the statements made by our professors of botany a few chapters back. Soluble Ca measured in the seed averaged over the 3 experiments had a weigh of 0.033 mg. The average weight for each plant was of 0.0511 mg or an increase in Ca amounting to 55%.
It is appropriate to state here the amount of Ca which passed from the roots into the water of the cultivation medium. I also requested an analysis of the water. Per plant this amount was of 0.0051 mg, or approximately 1/10 the amount of the content in the plant.
To summarize:
Ca: In Seed ~ In Plant
Ca insoluble: 0.0032 ~ 0.0319
Ca soluble: 0.0350 ~ 0.0511
Ca in water at end of experiment: --- ~ 0.0051
Total: 0.0382 ~ 0.0881Or an increase of 152% in total Ca.
The comparisons can only be approximate because the measurement of Ca was only performed by chemical analysis on the ashes. However, the direction the variations are taking remains the same. The increase in soluble Ca from the seed to the seedling should also be noted. It is 55%. The data given by CNRS shows a corresponding increase of 52% after mass spectrometer analysis and standardization to 100 gr of dry material in both seeds and seedlings. I was not briefed by CNRS about their procedure for preparing the material prior to analysis by the spectrometer. I do not know if these measurements were performed directly on a powder, on a spark or on a solution. In any case, the two figures are very close (52 vs 55%). In both cases there is a clear increase in Ca, which could not possibly result from an underestimation of the total Ca in the seed. On the contrary, if some techniques lend themselves to underestimation, they concern the seedlings and not the seeds.
Conclusion for this Chapter ~
]
I believe I have shown here that some chemists were conceited to the point of being laughable when they systematically, out of misplaced pride, rejected the findings of some of their colleagues. Why dont they themselves perform repeatable experiments with verifiable quantified data? Unfortunately, many people tend to deny the validity of other peoples work. They alone can detect the shortcomings of their peers; they carefully avoid doing anything, so they do not run the risk of being wrong. It is so much easier. This is not science anymore; it is malicious polemics.I believe that the scientists mentioned here, all highly respectable in their profession, will accept that their research quoted in this document, will be checked. They all agree that analyses of various major elements show completely different results for the seed and the plants under strictly controlled conditions. Water used for the cultivation is twice distilled; the air is extra-pure. No contaminating mineral salts may be brought from the outside. They observed the facts experimentally. They gave quantified data. But they did not interpret their data, in the hopes of remaining totally objective.
It is our role to interpret them.
I did not mention empirical observations in industry, which were brought to my attention in 1960, because I wanted to dedicate the first chapters of this book to the increase in Ca in oats during germination. The curing of potting clay by potters has been practiced since antiquity. It is due to fermentation. So are the malting and the fermentation processes in breweries. How many more examples couldnt we mention!
Part II
Explanation Of The Phenomena By Modern Physics ~ Theoretical Study
Chapter 1 ~ Explanations From The Atomic Particle Theory
Chapter 2 ~ A Few Examples Of Theories Proposed By Physicists
(1) Process developed by a US Army scientific department ~ (2) L. Romanis contribution ~ (3) Theory proposed by A. Dubrov ~ (4) A few interesting points of view expressed by physicists ~ (5) O. Costa de Beauregards theoryChapter 3 ~ Addenda To The Theoretical Study Of Low Energy Interactions Applied To Particular Transmutations
(1) Effective section ~ (2) Isotopic variations ~ (3) Nature operates at a finer level than man ~ (4) Changes in our understanding of some aspects of physics and weakness of this understanding ~ (5) A few arguments against a unified theory for electromagnetic and weak interactions ~ (6) Intermediary vector bosons
"Felix qui potuit rerum conoscere causas ~ Fortunate is the man who succeeds in knowing the (secret) causes of things" ~ Virgil (Georgics)
Chapter 1
Explanations From The Atomic Particle Theory
In order to explain a phenomenon, one must place it in a framework accepted by at least the majority of scientists.
After I established the existence of biological transmutations, I suggested global formulae at the atomic level. This was an entirely new idea at the time. It was then impossible for me to place these formulas in the frame of the strong interactions of the atomic bomb type, which was generally accepted. I was sure of the experimental results but these results could not be explained by the prevailing atomic physics of the time. I did not challenge this well-proven discipline in the least. As I had such strong evidence, I stated instead that we were dealing with a completely different phenomenon outside the realm of strong interactions. I had no accepted theory to propose, but I firmly stated that the strong force theory did not apply to the cases at hand and that it was up to the physicists to find another theory.
Theories are built on facts and not the opposite, especially when it is obvious that specific phenomena cannot be explained by these theories. Too many physicists keep in forgetting it. I had the opportunity of mentioning some exceptions to this unfortunate behavior; among the exceptions are the respected scientists: O. Costa de Beauregard, R. de Puymorin, L. Romani, J. Barry and others in France. In Russia I referred to Dubrov, Nejman and Korolkov in my previous works; several others wrote to me on this subject. In the USA, there were Dudley, Myers, Maxey, Bird and others. In Japan, there were Sakurazawa, Odagiri, Maruyama and others. Zundel and a few others, such as J. Boucher, performed experiments and did not formulate any theories. This was also the case in the USA, Argentina, Canada, and India as well as in other countries.
As there was no theory to explain these phenomena, I could only offer analogs to show that various structures of the components of the atomic nucleus were possible. I did this over a period of years. I showed that some schematic explanations, in fact simple vulgarizations based on the average energy per nucleus concept, were only figments of the mind. I showed that they could not be verified and that they were certainly wrong. There was room for other analogs.
An analog necessarily gives a wrong view of a phenomenon. It leads to representing actions at the atomic or subatomic levels as if they were working at the level of our senses or of the cell. In fact, these actions cannot be detected at the molecular level, even less by our senses.
These considerations are only secondary, because the general public is primarily interested in applications. Fortunately for our successors, we will leave them much to discover. As important as application may be, it is not everything. There is also the satisfaction of knowing, the interest found in abstract knowledge, which is a strong intellectual stimulus.
We had to wait 15 years, until 1974, before theoretical physics brought us the elements which were formerly beyond our reach. The readers, who read the previous work I published in 1975, may have noted on page 281, that on July 23, 1874, I wrote to the physicist O. Costa de Beauregard to confirm the existence of neutral currents. These currents were first discovered in Weinberg and Salams theory. I believed that this theory could provide a base for the explanation of transmutations at low energies. This supported the role of neutrinos in the reactions I had proposed, a role I had suspected. By the second half of 1974 de Beauregard had succeeded in refining the Weinberg-Salam theory and adapting it to my work. This adaptation was included as an epilogue to my book, dated December 1974 which was already at the printer. Dare say that shortly after that, in January 1975, a Nobel Prize official delegate nominated me for the prize in Physiology and Medicine. The physiologists did not want to take precedence over the physicists. Weinberg received his Nobel Prize in Physics only in 1979. This confirmed that this theory, the one I mentioned in 1974, was valid and internationally accepted.
Those who have little interest in theoretical considerations may choose not to read Part II of this book, but I believe that many will try to understand it. I will attempt to be understood by the general scientific public; only the final chapter will be written for nuclear physicists. I will keep in mind that for many of these, this is a completely new and unknown subject. Due to their ever-increasing specialization, nuclear physicists tend to limit themselves to the physics of the strong force and to focus only on some parts of this widely expanding science. For them, this will be an introduction to the physics of weak interactions, a subject which is still little known, especially in this form. This branch of physics is too new to be the subject of many high quality publications. There is always a lag of several years between discoveries, the synthesis of the new information they brought, and the acceptance of this new information by the majority of the theoreticians. When I started planning for this book in late 1978, I thought I would complete it in 1979, and at that time no works on the physics of the weak interactions were available. I decided then to include a chapter on this subject. It is indeed somewhat abstract, but I believe it will be useful to the average scientist unfamiliar with this branch of physics. The situation evolved after the award of the 1979 Nobel Prize in Physics. The subject is now better and more commonly known; I decided to modify the plan of this book. It is impossible to treat a subject in full evolution so the book remains up-to-date for several years. Therefore, I thought that the scope should be modest and kept general so it would not be contradicted by the discoveries in process. These discoveries fundamentally change the picture presented below.
I will not mention strong interactions, which does not mean that I reject them. This would be silly on my part. However, it should be recognized that too many physicists and too many of their followers, scientists of various backgrounds, tend to grant too wide a scope to the data in nuclear physics. One can always argue about an extrapolation and sometimes an extrapolation may be plainly wrong.
I do not wish to argue about Einsteins laws. I accept them, although I do not always agree with the people who use them any old way. The postulates, on which they were based, should always be kept in mind. Indeed, Einstein never extrapolated these laws. E reminded his dogmatic worshippers about their limits. In particular, he noted that his laws did not apply to biology. He could not say why, but it was obvious that biological experiments were not ruled by his laws. Einstein did not know about the neutrino when he formulated his laws. He only knew that his knowledge was limited and that a lot of nonsense had been spoken and taught about the restricted relativity law E = MC2. In this law C2 is a constant and M is proportional to E (and vice versa). It always remains a linear law, a first degree curve; in other words, a straight line. This is why he had to search for his General Relativity Law, in order to explain the curvature of light in the vicinity of large masses such as the sun or any large star. Curvature means change of velocity, a factor of the second order. So many scientists forget that M is not the stationary mass designated as Mo; it is in fact a relativistic mass.
One often forgets also that the mass-energy conversion is not a simplistic rule. We do not know how to convert any old mass to energy. We first have to use some energy which produces anti-matter which in turn gives energy after annihilation. We do not know how to destroy a proton or a neutron. These particles remain unchanged in an atomic explosion or in a fission reaction. Only a small part of the linking energy between nuclei is used. I do not intend to discuss the heavy particles, the baryons, or their components, such as the quarks, or even the gluons which supposedly link the quarks. There are vast problems in this area which is in full evolution, but these problems do not concern the fields of weak energy. Indeed, we will mention the hadrons in our study of weak interactions. The first statement on the existence of weak interactions was made soon after the discovery of the neutron. Obviously, a neutron was not simply a proton plus an electron. There was a particle, which could not be detected at the time, but which was necessary to restore mass balance. It was named neutrino. A very careful study of natural radioactivity showed that there was an electromagnetic phenomenon, the emission of one electron, but that there was also the emission of one anti-neutrino. Similarly, we will introduce the notion of proton movement in the course of our study of weak interactions, in relation with other phenomena such as the tunnel effect or the effective section.
We will study these phenomena a little later. Other particles must also be introduced. They are necessary to the theory, but they have not been experimentally isolated yet. These are some of the essential elements of the theory, for which the 1979 Nobel Prize was awarded. They are the intermediary virtual vector bosons. We will spend a little more time on them.
As early as July1974 I had focused on Weinbergs theory, which was followed shortly by Salams theory. I related this fact in a letter excerpt in the book given to the printer at the end of 1974 and published in early 1975. I applied this theory to a biological phenomenon I had discovered. But what is more important? The proof of the existence of a biological phenomenon or the theory behind it? It is not up to me to judge. I cannot be the judge and party at the same time. Furthermore, how should we judge when the problem straddles across biology and physics, and when specialists of either discipline will be called upon to cast a judgment. Each one tends to form an opinion on the basis of his own knowledge. It is only human. There are no interdisciplinary Nobel Prizes. There is a Prize in Physics, another in Physiology or Medicine. Specialization is a rule everywhere, especially in physics. The biologist does not want to innovate and he follows the physicist. This is why the official delegate who nominated me for the 1975 Nobel Prize I Physiology or Medicine was not followed by his colleagues on the Nominating Committee. He was not a Frenchman. Nobody is a prophet in his own backyard. Only in 1979 did the Physics Nominating Committee award their prize for the Weinberg-Salam theory which I had adopted 5 years earlier. Lets not dwell on the past. The essential was to prove that a fact scientifically established after many indisputable experiments was in line with the classic theory. I am trying to show here that the clear evidence obtained for this phenomenon and its multiple implications, represent a major turning point for Science internationally. Theories other than Weinbergs were proposed throughout the world to explain the phenomenon. I believe that the most widely accepted theory was proposed by the physicist de Beauregard. It was refined during the second half of 1974 and its principle was then accepted by Bernard dEspagnet, a well-known atomic particle specialist. The theory was summarized in my book of 1975. It was not refuted, but it was only a schematic explanation. I completed this theory by introducing the intermediary vector bosons in various publications as early as 1976. This is the reason why I will give more attention to this theory and I will describe only briefly a few others, which incidentally provide very interesting complements to this theory. I will describe this complementary information in greater or lesser detail.
Chapter 2 A Few Examples of Theories Proposed by Physicists ~
(1) Process Proposed by a US Army Scientific Department ~
A department of the US Army Scientific and Technical Services headed by S. Goldfein studied my research. The team gathered for this purpose worked on the report from December 1977 to April 1978, and the report was published in May 1978. It was distributed to numerous specialized service in the various branches of the Armed Forces (Army, Navy, Air Force, General Services). It included 28 pages, not counting the cover, and 8 figures. In it Goldfein proposed an explanation based on physical chemistry, delineating a possible process which must still be developed and defined more precisely.
Numerous calculations made by the authors of the report, too few according to them, showed homogeneous results. They showed that the reactions originating from my research confirmed a gain in energy. There was a new source of energy, which had to be investigated more closely. In our time no source of energy should be neglected just because it was insufficiently studied. This was the reason this agency, highly respected worldwide, decided to intervene.
Classic physics shows that K + H >> Ca + 0.008 a.m.u. This means that in the reaction v + H + K >> Ca + v, the energy taken by the outgoing neutrino v should be greater than the energy brought in by the incoming neutrino v. In other words, in biology the production of energy should not be limited to the sole exoenergetic chemical reactions. Exoenergetic reactions of physical origin should also be considered, although the energies involved are modest compared to the energy of fusion by the strong interaction. They are far from being negligible, as they result in a positive balance of approximately 40% for the cases studied.
We noted from de Beauregard: v + p >> v + p with v =/ v (see the last chapter in our book of 1975). The US Army Scientific Services state: v >> v and v ~ v + 40%. Could this be attributed to a fusion process produced following a process other than the strong interactions process?
The report describes an example based on the oxidative phosphorylation process in mitochondria. Some animal cells include up to 7,000 mitochondria. From the energetic point of view, the active molecule is ATP after chelation of one Mg atom (Mg-ATP) under the action of an enzyme, Mg-ATPase. I discussed this process in more details in my book of 1975, in particular on page 85. I first presented the beginning of its study in my book of 1968, now out of print.
The US Army study shows also an action by D-ribose molecules interacting with the Mg-ATP to produce a rotation of the acyl-oxygen dipoles linked to P atoms. Schematic representations and computations lead to a helix with H+ ions in an unsaturated ionic structure. A link is established between the D-ribose and the form gamma oxygen. The Mg++ chain electrons (axial chain of 10 Mg++ in the study) produce an oscillating electric field leading to a resonance. An H+ ion, introduced between the components of an OH and gamma-O pair, ends up on a circular helix trajectory of approximately 30 Angstroms diameter under the dipoles impulsion. Following this hypothesis, the hydrogen H+ positive ions would finally acquire a very high rotational velocity. This would be a relativistic velocity, so to speak, due a cyclotron effect, such that H+ would cross the potential barrier of an atomic nucleus and penetrate into it. If the proton receiver is an atom of K the reaction would be: K19+ + H1 >> Ca20.
The development of this hypothesis fills about 14 pages of the report. It is very interesting because it provides an explanation for a physico-biological process partially established by various studies. This hypothesis should be applied to other cases. I thought it was opportune to mention it here because the US Army could not pursue the research for lack of funds, as it commonly happens everywhere in the world. At least the agency wanted to show the solid bases for the hypothesis.
In the first part of this chapter I did not intend to give my full and unconditional approval to the study. The author of the report was fully conscious that complementary studies were necessary and that the study completed constituted only a first step. He would have liked to receive sufficient funds to be able to take one more step. At the time I am writing these lines, this has still not occurred.
It is obvious, for example, that special procedures should be applied for the measurements related to energy balances. These measurements are very complicated. Known calorimetric methods cannot be applied, because neutrinos intervene, as we are here in the domain of weak interactions. Neutrinos interact only very seldom with matter, A large part of the energy is carried away under a virtual form, so to speak. It goes through space without affecting our senses or our measurement instruments. How can we measure it?
How can we measure the energetic contributions from enzymes or from ATP? Here we are dealing at the molecular level. There are uncertainties in the values measured indirectly by methods based on theories which may be questioned in the future.
Many more points need to be investigated more thoroughly. Much uncertainty remains regarding the quantitative study of energy balances and also regarding the qualification of the operator. Many questions should also be raised on various theoretical points. It is pointless to list them here. The main point is the experimental study leading to the confirmation or the refutation of a theory. It is not the blind respect of theory.
I congratulate S. Goldfein for his report. Synthesis of physics and biology, such as the one he presented, are rate. There is here a wide filed for research which is not even being explored in most countries. Some aspects of the physics were left aside by the US Army scientists, weak energy interactions in particular. Few traditional physicists knew about the work of Weinberg and Salam before it was widely publicized after the Nobel Prize award at the end of 1979. S. Goldfein and his associates stated some synthetic considerations of the highest interest and their study is very encouraging. I have condensed too much the published report any my summary may be inaccurate, because incomplete. The subject is so vast that it is difficult to condense. Some American magazines presented summaries even more succinct than mine in order to prod the reader to refer to the original report. The reputation of the agency which issued the report is a guarantee that the study was done in earnest. My research reached renowned agencies throughout the world. Its value was confirmed by my nomination for the Nobel Prize by a scientist belonging to this internationally known agency. I started my presentation with the US Army study, because to my knowledge, it was the most detailed study published by an author and because the synthesis of physics and biology is most interesting despite the reservations made by theoreticians on some of its pints. The future alone will tell.
(2) L. Romanis Contribution ~
I must also discuss at some length a study made by the physicist L. Romani because it was not published insofar as I know. L. Romani is a well-known fluid mechanics specialist. He is known in particular for his remarkable contribution to the original, simple and efficient solution of specific technical problems in the field of wind energy (automatically controlled windmills) and of wind effect prevention (on suspended bridges, for example). He is the director of the Eiffel Laboratory, in which air flow studies are made for the purpose of improving aerodynamic shapes (aircrafts, automobiles, etc.). Eiffels laboratory facilities include a wind tunnel, which is used to check computations on scaled models. This work brought L. Romani to reexamine some aspects of acoustical waves and more generally of all waves.
His keen understanding of energy problems related to air flow brought hi to study my work because of its implications in the field of energy. As soon as 1963, he proposed for it a purely physical examination which I presented in part in a book published in 1964. The validity of his calculations was accepted by two respected scientists mentioned in his paper. However, it seemed that the generalization of the phenomenon ran against some observations pointed to me by a well-known specialist of wave mechanics. For this reason I did not publish the remaining of Romanis study in my subsequent publications. I did not even include the text published in 1964, as this study presented only a historical interest.
However, Romani did not forget the matter. To this day he was not able to propose a global explanation of the process encompassing all the points established in my research. Nevertheless he showed that one should not forget particular fundamental points resulting from the study of wave phenomena if one does not want to become lost in farfetched theories.
This respected physicist published a basic work in two volumes on his theories, Theorie Generale de lUnivers Physical... [blurred line in photocopy]... (A. Blanchard, Paris). He frequently used dimensional equations. As far as my work is concerned, one should refer mainly to partial studies, in particular to mimeographed copies of lectures he gave at the Sorbonne and in various other places mainly from 1976 to 1978.
In November 1976, for example, he expounded various original points of view on energies at Cercle de Physique A. Dufour in relation to Relativity and energy transmission via the ether, which he still fully supports. He defended his position on this subject in multiple lectures. He accepts four types of energy, as everybody does today in a non-definitive and non-limitative way. He also stated the following:
(1) The body on which the gravitational field is exerted, or mass, has the dimension of an inverse length L-1;
(2) The body on which is exerted the weak nuclear field, or hypermass, has the dimension of a Gauss curvature L-2;
(3) The body on which the electric field is exerted, or charge, has the same dimension as the ratio of a torsion by a curvature. It is a pseudo-scalar number;
(4) The body on which the strong nuclear field is exerted has te same dimension as a torsion, L-1.
One can see here some of the fundamental differences in the nature of the fields for the four types of energy. These lead us to express some reservations on some unified theories which can only unify everything in confusion or in a generalization devoid of precise meaning.
I will come back to some of the differences in the main characteristics of the representative waves for these forms of energy, as they were outlined by L. Romani. I will only mention in passing various pertinent comments made by L. Romani, which are forgotten too often. According to him, photons are not waves but they are pairs of whirls. This concept leads him to original views which are far from being unanimously accepted by physicists. Waves do not carry energy; they carry information. Romani is far from admitting the classic definition of information as given by Brillouin and others. To consider information as the co-logarithm of entropy would be a mistake, as information cannot consume any energy. Otherwise the French language would not make sense anymore! But is it all that new? Blaise Pascal wrote: "I never argue about a name, provided I am told about the meaning which is given to it". Dialectic is still with us. A piece of information is a data, something abstract; it is something potential. It is only when the information is used that there is energy consumption. The whole entropy question should be reviewed, as it is generally agreed. In 1977-1978, J. Tonnelat addressed this problem in a work in two volumes. He showed how poorly defined the problem was. This problem is extensively reconsidered in J. Tonnelats books. The reader will have a different view of the entropy (and negentropy) problem after reading them. "It is not right to consider entropy as energy, although both are dimensionally equivalent" (p. 158). One should also keep in mind some considerations which the author does not even mention. In particular, and I insist on this point, the problem of energy in living matter should not be looked at in the sole light of thermodynamics. Lets not forget the action of neutrinos coming from the ambient medium (lets say cosmic; lets not be fooled by words) on the life phenomenon. This is an essential point for our work, as we will see later.
Lets come back to a remark by L. Romani: a wave only propagates a movement. The associated photon (electromagnetic energy) carries the information. In each point, at every instant the kinetic energy of the undulatory movement is derived from the potential of the Ether. There would be no energy transport by the waves of the local Ether. These waves are used as signals; they transport the information. Information initiates a similar movement in the receiver, which derives the necessary energy from the Ether and transforms this energy into kinetic energy, the only form of energy we can perceive. This is why a wave appears to carry energy. Romanis Ether concept is new; it is the "tense" Ether.
(a) Waves and Low Energy Transmutations ~
I will not summarize here L. Romanis views on waves. He dedicated to them Ondes Inconnues (which was only mimeographed), a very dense work, too long to be summarized. However I will refer to it in the later part of this book, because Romanis views on waves are too important to be ignored or forgotten. It is essential to keep them in mind in order to remember the fundamental differences between the various manifestations by other forms of energy fields. For the time being, I will only quote a few statements made by Romani:
(1) Strong nuclear energy field waves are either centripetal or transversal and circularly polarized;
(2) Weak nuclear field waves are longitudinal and circularly polarized.
I emphasize that only circularly polarized energy forms, as defined by Romani, seem to be capable of producing element transmutations. This is a personal comment and it should not be attributed to Romani. I am only borrowing his wave classification, not the effects of the waves, which he did not mention. Wave effects were derived form my studies. I do not pretend to establish a general and absolute rule, valid for all transmutations, but only for the specific transmutations I was able to check.
Gravitational waves (if there was such a thing) would be longitudinal and not polarized.
Electromagnetic waves would be transversal with a rectilinear polarization.
It is often said to simplify that "all simple displacements are either rotations or translations. Furthermore, a wave is either transversal or longitudinal". In short, it follows that:
(1) Gravitational waves are longitudinal and not polarized;
(2) Weak field waves are longitudinal and circularly polarized;
(3) Electromagnetic waves are transverse and linearly polarized (in a plane perpendicular to the direction of propagation);
(4) Strong field waves are transverse and circularly polarized.
The first two longitudinal types can pass through shields, just like sound waves.
In fact, undulatory phenomena are seldom simple. In the course of their propagation, waves may differ from the original forms in which they were emitted. Transverse waves may be linearly polarized in the plane perpendicular to the direction of propagation. Hybrid waves may occur such s waves polarized in a plane including the direction of propagation. Others are transverse and circularly polarized in the plane perpendicular to the direction of propagation. They could consist of the sum of two conjugated waves in quadrature.
The wave question is complex and I cannot suggest any work covering the whole subject. The course by J. Bo and N. Hulin-Jong (Hermann), Ondes Electromagnetiques-Relativite, brings up interesting points of view, but it tackles only one type of wave and does not provide for any comparison, as it deals only with electromagnetic waves. These waves cannot cause weak energy transmutations, which are associated to waves of a completely different nature.
I will come back later on specific ideas of Romani which I associated with some effects which he did not mention. These effects are in line with the additional information that Romani sent to me in May 1977 and that I discussed at the ARK-ALL inter-university meeting of June 1977. My paper was printed in ARK-ALL Publication 1978, Vol. 4 (1): 49-63. I will have the opportunity to come back later to some fundamental differences between weak energy fields and other types of fields,
(2) Theory Proposed by Dubrov ~
In various letters, Alex. Dubrov, Dr in Science and Member of the Geophysics Institute of the Sciences Academy of Moscow, shared with me his ideas on the theoretical mechanism of biological transmutations. I will give here a brief summary of the information contained in The Geomagnetic Field and Life, a book he published in the USA in 1978. This was a corrected and much enlarged version of the book in Russian published in 1976. Dubrov sent me a copy of his initial book in Russian.
The Geomagnetic Field and Life-Magnetobiology is a big book published by Plenum Press, NY. It contains a bibliography of over 50 pages as well as 82 figures. In this work the author gives many experimental results related to electromagnetic effects of the earth on live matter. Some of the points studied were accepted by Prof G. Piccardi, formerly Director of Physics Laboratory of the University of Florence. I met Prof Piccardi several times and he sent me his book The Chemical Basis of Medical Climatology, published in the US in 1962 by C.C. Thomas, Springfield. He spent much time studying the effect of solar fields on the rate of precipitation of colloids, on human colloids, cells and blood in particular. He published Biolelectric Rhythms in Human Blood in Holland, 1970.
Dubrov believes that geomagnetic effects are exerted at a very low level, even at a subatomic level, in the framework of the weak energy interactions. Geomagnetic field changes could reverse the spin of elementary particles, transforming a [ 1 sentence missing text ]. In some cases they could cause dissymmetry by mirror effect (a 180 degree rotation). These molecular modifications in turn could alter the oscillatory rhythms in several parameters of the living organisms. The infractions to the symmetry law are important. They remain very much in the actuality, as attested by the 1980 Nobel Prize in Physics. This prize was awarded to a physicist who researched some specific infractions to the law of symmetry. His work was done some time ago, but the award shows how much interest the infractions to the law of symmetry still arouse.
In 1980 Dubrov published another book in Russian on mirror symmetries. He sent it to me, but I do not know if it will be published in English. He advised me of the publication by Planum, NY in 1981. This new book will be co-authored by an America named Pushkin. In his book of 1978, Dubrov applies the geomagnetic field to biological transmutations, following the formulas I proposed several years before. He mentioned these formulae in his book of 1980. It seems that he did it from memory as he made some errors.
He quoted some studies which show that magnetic fields of 1,000-5,00 oersteds modify the amounts of dry material and ashes in plants as compared to seeds from which they originated. No minerals were brought from the outside during these studies. The same fields cause changes in oligo-elements according to these studies, which could only be explained by a transmutation of some elements.
The cumulative effect of changes in the earth magnetic field, due to earth rotation, of proton showers and other factors, could produce structural inter-chromosome inversion by rotating one chromosome segment. In so doing it would modify the coding for the synthesis of enzymatic proteins. According to Dubrov, the genetic code would be based on a pentametric symmetry and the RNA triads would be arranged in an icosahedron. If these triads would be arranged in an icosahedron. If these triads were rotated by 180 degrees, alterations in residual amino acids could result. A change in the position of a single asymmetric atom is sufficient to modify the property of elementary particles which are becoming symmetric. In this fashion racemic tryptophan could be transformed into laevo-tryptophan. Lets remember the importance of asymmetric carbon in organic chemistry.
A magnetic spin reversal could be the initial cause of this transformation according to Dubrov. Elementary particles are very sensitive to variations in the parameters which involve weak energies. In this way there would be a correlation between macrocosm and microcosm.
Geomagnetic field effects should be studied more closely in the light of the progress made I wave mechanics, communications, cosmology, and geophysics. They show tat there is no theoretical impossibility in accepting that some biological transmutation do not belong to the domain of strong interactions and that they can be explained by weak energy interactions.
Dubrovs study is most interesting because it emphasizes the mechanism ... [missing page in text --- badly blurred photocopy]
According to Dubrov, there could be no significant interactions between neutrinos and living matter without actions by the enzymes, whatever the direct cosmic effects. As member of an agency affiliated to the University of Moscow, Dubrov focuses on a point worthy of additional studies. For the time being he shows mainly the directions which may lead to the explanation of the phenomena. He barely touched on their quantitative study.
(4) A Few Interesting Points of View Expressed by Physicists ~
Various theories were proposed to explain the phenomenon of transmutation by living matter.
The physicist Rene de Puymorin thought of a stationary wave mechanism similar to the sound mechanism. Based on the computation of several specific data, he concluded that electrostatic repulsion was wavelike and that it was not continuous or hyperbolic. He states its wavelength and from there his computations enable him to determine other aspects of the potential barrier. His study is primarily based on a specific concept of the electron structure. I am afraid of distorting his point of view in summarizing it too succinctly. The reader should refer directly to the work LOrigine de la Gravitation, published in 1975 by La Pensee Universelle, Paris. In this book R. de Puymorin expresses different views on various problems presented in a fairly simple and very personal way. It is food for thought. This small book of 64 pages is easy to understand. Puymorin discusses biological transmutations in 11 pages only. He sent me several other more extensive studies which had never been published. I cannot refer the reader to them, and I have no room here to include the various notes puymorin sent me.
(5) Costa de Beauregards Theory ~
As early as 1963 the physicist O. Costa de Beauregard saw the possibility of introducing the action of neutrinos in the reactions which I published in 1960. I noted this possibility in my book Transmutations a Faibles Energies, published by Maloine in 1964. C. de Beauregard said he was convinced that the energy balance in my reactions could only be understood though the effect of massless and chargeless particles.
He immediately ruled out neutrinos, of which he thought first, because the conservation principles were not respected. Only 10 years later could he formulate a theory satisfying these objections, at the time the neutral current theory was introduced. This theory constituted a major progress in the physics of elementary particles.
This theory was summarized by the simple formula: p + v >> p + v , or: iN + p1>> pN + v .
I had observed that such reactions were experimentally verified only for odd A nuclei and that they yielded modified even A nuclei with a 1H1 (or 1p1) proton. At the time none of my experiments showed that it was possible to jump from an even A to the A of rank immediately superior. No experiment showed either that it was possible to go down scale from N +1 to N by a single step.
We had: p =/ p and v =/ v because the energy of the incident p proton was different from the energy of the p proton included in the nucleus. A neutrino of v energy was absorbed and a neutrino of v energy was re-emitted. The energy difference was emitted into the ambient space without thermal effect or without any other effect on the material. This explained the energy balance observed. Such a formula satisfied the most rigorous conservation of energy, mass, impulse, angular momentum, spins, baryonic and leptonic numbers, etc. It could not be refuted with any theory.
I will not give more details on de Beauregards theory. He presented it himself in the epilogue of my book, Preuves en Biologie de Transmutations a Faible Energie (Maloine, 1975). In the last chapter I reported on our active correspondence during the time de Beauregard was working on his theory. The principles of this theory came to his mind in the spring of 1974 after the existence of neutral currents was confirmed. He asked me to insert it at the end of my book, which was then at the galleys for publication in 1975.
A neutrino provided the necessary impulsion so that a proton can be included in an atomic nucleus by tunnel effect in accordance with quantum mechanics. This was the basis of de Beauregards theory. For example:
V + K19 + H1 --> Ca20 + v1 ( with v =/ v )
The v neutrino is expelled in order to reestablish the energy balance between the two sides of the formula. The incident neutrino v provides the initial energy or the impulse which permits to introduce the H+ into the nucleus of the potassium atom.
Costa de Beauregard noted that his theory was only a preliminary framework and that it had to be completed. To this end a change in the transition natural possibilities under the effect of an information structure inherent in the life phenomenon, had to be considered, according to him.
The theory still holds after 7 years if progress in the physics of elementary particles. As soon as 1976 I introduced various complementary concepts to enlarge and further document some theoretical physical aspects of the basic principle which was published in early 1975. These complements engage my sole responsibility. They cannot be attributed to de Beauregard I any way.
Chapter 3Complements To The Theoretical Study Of Weak Energy Interactions Applied To Some Transmutations
As soon as 1975 I studied some complementary ideas which I thought necessary in order to enlarge the theoretical physics basis laid down by de Beauregard.
Both of us lectured in London in 1975 in front of a large audience during the May Lectures and the morning after in front of specialists at the Kings College.
These developments were the subject of various publications in French and English from 1976 through 1978, especially during the latter year. Different aspects of physics and progresses realized in this science since 1975 led us to define more precisely the initial concepts. I will briefly describe here the scope of our knowledge as of the beginning of 1981. Fundamental concepts are too often unknown of the physicists bogged in obsolete ideas. These concepts have been insufficiently publicized, unfortunately. This explains why these physicists are taking positions incompatible with the new science of the weak energies interactions. I can only discuss here a few fundamental points.
An objection, still raised in 1980, is that the effective section of interaction between neutrinos and protons is too small to permit a weight change in the material. O. Costa de Beauregard had precluded this objection in his ending note to my book of 1975. At the time he based his concept of effective section only on classical theoretical considerations accepted by most physicists.
Already in 1974 this notion was contested. It was based on the results of statistical evaluations which were too general, oversimplified, sometimes purely imagined and not confirmed by precise calculations. Incidentally, one did not know how to perform these calculations at the time. The question was more complicated than it was assumed at the time, and in a sense it was premature. The subject was discussed in lectures given during the 1974 summer session at Gif-sur-Yvette. In the June 1974 of Physics Today, it was noted that the neutrino-hadron effective section did not have a fixed value and that it did not vary only with the energy of the incident neutrino. It was proportional to the square of the atomic mass of the nucleus. On other words, if the target was a K39 nucleus, then the square of this number should be introduced in the calculations. A considerable scale change resulted. A neutrino, which passes through a nucleus, comes close enough to all the nucleons in the atoms to react with each one. It is not absurd to conceive that the electrons other than the K orbit, revolve around the concentrated group of nuclei, when nuclei are very close to each other. This is the case in molecules, in which nuclei keep only around themselves the two electrons revolving on the K orbit. These electrons can even revolve in the interspace between nuclei. In this way there are many different orbits. Some organic molecules may have molecular masses reaching several tens of thousands, even hundreds of thousands. The effect of an incident neutrino is felt by the whole nuclei cluster.
Additional studies were going to support my ideas. In a 2-page paper published October 6, 1975, in Physics Letters entitled "Are Solar Neutrinos Detected by Living Things?", Ruderfer quoted the results of experiments performed on the giant calmar. He computed the effects of these neutrinos on mammal brains, as he had noticed that the sensitive part of a neuron (axons and dendrites) can amount to 100-1,000 times the volume of a body cell. This brings us to review the notion of capturing section, or more precisely, of effective interactive section. A 5-page study in English by Ruderfer, "Neutrino Structure of the Ether", published in Lettre al Nuovo Cimento (May 3, 1975), makes for an interesting read. So do other articles on neutrinos published by the same author in this periodical as well as in other American magazines. Some of these articles date back to 1968 and they deal with physiological and psychic points of view.
(2) Isotopic Variations ~
I mentioned several times the wrong postulates accepted as classic by some physicists and phytobiologists. These errors are taught in schools of science, in schools of agronomy of high level, etc. Among these the invariance of the isotopic composition for a given element is most commonly accepted.
I pointed out for example that photosynthesis is not a uniform phenomenon for all plants. His was a recall for some and a first for others.
Isotopic variations explain the discrepancies in atomic mass between studied vegetal samples given by analysis. In my first book, published in 1962, I quoted discrepancies in the atomic mass of K measured with a mass spectrometer in potatoes. The mass varied according to the variety. These variations were not mentioned because they were attributed to errors in the laboratory procedure. Any other explanation could not be believed.
Too many physicists and in their lead, phytobiologists, chemical physicists, medical doctors, agronomists and others, still ignored at this date these ideas seldom taught in school. For them, the isotopic composition is stable and the atomic mass is given in chemical tables.
For how many physicists, even nuclear physicists, this axiom has become a taboo. Some told me, "Your reactions will only be accepted when you show that initial isotopes correspond quantitatively to final isotopes". To illustrate their point, according to them the increase in Ca in a plant must represent the sum of an atom of K and an atom of H, such that K39 + H1 >> Ca40 or K41 + H1 >> Ca42. There is very little heavy hydrogen (deuterium) and it can be neglected usually. However it is not always the case. It is obvious that all the stable isotopes of Ca cannot be derived from the stable isotopes of K and H. I will state again that so far I never observed that Nature had to start from radioactive isotopes and that it could not produce radioactive elements. I do not pretend that it is not so and I insist on this point: I never observed it personally. Studies were made on this subject with extremely sensitive instruments, as at the French Centre dEtudes Nucleaires in Saclay. Hence it was proved that the N15 + O16 >> P31 reaction by microbial action could only be obtained from N15 and O16, and that all other P isotopes would be unstable and radioactive.
All radioactive isotopes found in living matter come from the outside, as it is the case in particular for C14 and K40; furthermore they are found there in microscopic doses. A complete study of these cases must still be made as they occur during germination. I lacked the time and the financial resources required for such a study.
Studies made on K and Ca isotopes show that the isotopic composition of a plant is different from the isotopic composition of a seed from which it originated. Photosynthesis should be kept in mind. We saw that it may vary according to the photosynthesis cycle specific to the vegetal family. This is why we can state that the people, who misinterpret the quantitative inequalities between isotopes in biological transmutations, make unwarranted generalizations. This should not be construed as a weak point in my observations, but only as the proof of their ignorance.
One should also keeping mind what I stated in my earlier books and what I summarized in my book of 1975. All the Ca does not come from an agglomeration, from a fusion of K and H. there are other sources which vary according to the calcicole or calcifuge character of the plant. Specific species may need an alkaline, acid or neutral soil. A legume is not cultivated as rye grass or oats. Some plant families can transform manganese into iron. The reaction may be reversed in other plants and in other soils. The reaction is as follows: Mn55 + H1 >> Fe56. In animals the following reactions occur frequently Mg24 + O16 >> Ca40. The various stable isotopes of Mg and O correspond to stable Ca isotopes. Other stable Ca isotopes may lead to think that they come from stable silicon and carbon isotopes. I dedicated several chapters to these transmutations in my earlier books. I noted that in 1799 Vauquelin suspected such a correlation, although he did not mention it. Nuclear questions were unknown at the time. His chemical analyses showed an increase in Ca and a decrease in Si which were not quantitatively equivalent. He could not have thought of the K + H transmutation. I did not have the time to investigate the various potential origins of the Ca. Simplifications are out of the question and generalizations even more so. Naturally Vauquelin could not introduce isotopes in his comparisons, as the isotope concept was totally unknown at the time.
I also showed the isotopic correlation Fe56 - He4 >> Cr52 which is valid for the four stable isotopes of Fe and Cr. He4 is the alpha particle. The correlation holds also to the few thousandths for the ratios between stable isotopes. I discussed this point in particular in my last two books. One should keep in mind this neutronic activation property. Its concept is completely ignored, even rejected with contempt, by many specialists. For them there are no differences in the compositions of live and dead matters. Life can only engage in chemistry, in which discipline nothing is lost and nothing is created. However, there is an indisputable variation during germination, as we showed in the first part of this book. We proved that the creation of Ca in oat after germination in a medium which could not receive any calcium from the outside. To compute the total Ca weight according to the isotopic ratio for a specific nucleus may seem to be a figment of the mind. However, one should not forget that the isotopic compositions given in the Atomic Mass Tables used in chemistry appeared to be so different from each other that it was necessary to define internationally the origin of the element selected as isotopic composition standard. Discrepancies are often greater than 1/1,000. In some cases they can be much greater, for the deuterium/light hydrogen ratio, for example, because the deuterium atomic mass is nearly the double of atomic mass of light hydrogen. In some reactions, decreases of up to 70% in H were noted compared to the initial isotopic mixture. This is the phenomenon which allowed isotopic enrichments by separation of light and heavy isotopes. This separation can be obtained kinematically by diffusion through the appropriately calibrated hole of a porous membrane. There is no transmutation in this case, but obviously one should have some reservation regarding some methods used to track Ca or other elements, by neutronic activation for example. This does not mean that this technique should be absolutely rejected from the outset, but that the results must be cross-checked whenever the analysis is performed on living material. By neutronic activation the stable Ca48 becomes radioactive Ca 49 which is easily identified with accuracy. It seems that the ratios between these two Ca isotopes and the total Ca remains constant in the seeds as well as in the plants. It is therefore possible to use them to measure the total Ca. However, this does not explain the origin of Ca42 in the total Ca in live materials. One should remember that there is only 0.18.1,000 (less than 0.3/1,000) of Ca48 in the standard. This extremely small concentration can only be marginally detected by many analytical techniques. This may be one reason why it is so difficult to trace the origin of this isotope. In oats its concentrations increase from the seed to the plant. Whatever happens, it is accepted that the Ca48/total Ca remains approximately constant. Total Ca variation is computed according to the variation in activated Ca.
In this regard the director of a neutronic activation analysis laboratory wrote to me in February 1976:
"We measure the Ca coming from the Ca48 >> Ca47 reaction. Therefore the activation analysis yields basically the amount of Ca. The total Ca is obtained by accepting (assuming) the natural distribution of isotopes".
For many physicists this acceptance is still an act of faith at the present day. There can be exceptions, though this is not obvious, as I and many others before me could observe it. However results obtained by the neutronic activation technique should never be accepted blindly; they should always be cross-checked by chemical analysis (a gravimetric technique in particular). Sometimes I have another cross-check made by atomic absorption spectrophotometry (with a Beckman or Perkin-Elmer instrument).
Macroscopically the question of the origin of the Ca isotope is unimportant, because this element represents only 2/1,000 of the total. However, I do not underestimate the theoretical importance it may represent and it is the reason why I discussed it here, so physicists would not forget, sometimes willfully, or would not ignore the biological causes of some physical phenomena, among them the isotopic selectivity of live materials.
(2) Nature Operates at a Finer Level than Man ~
How much progress was achieved in chemistry in the last 100 years! It is now to the point that chemistry invades everything, that all products on the market owe their existence to chemistry and very few products are natural anymore. Unfortunately there are people who do not understand that this ingestion of chemical products damages our bodies.
People who are for the use of chemical products pretend that natural and synthetic chemical products of the same formula have the same effects on the human body. They state that their effects on our body are necessarily the same, because they cannot be discriminated a=from the chemical point of view, and for them al of Life phenomena are chemistry.
It is their mistake. Chemical analysis is not enough to characterize a product. As a matter of fact, biochemists agree on this point now. They know that qualitatively and quantitatively identical atoms can be located in space in different ways, and that for this reason, resulting external electron envelopes may be different.
Translation ends here. The following sections are not available in English (or perhaps my photocopy was incomplete):
...(4) Changes in our understanding of some aspects of physics and weakness of this understanding ~ (5) A few arguments against a unified theory for electromagnetic and weak interactions ~ (6) Intermediary vector bosons
Appendix 1 ~ Geology
Appendix II ~ An Extrapolation: (1) Crying wolf ~ (2) One must rethink the concept of energy in living matter ~ (3) Non-electromagnetic energies in living matter ~ (4) Parapsychology and hypothalamus ~ (5) Germination procedure for oat seeds with no external addition of calcium
Appendix III ~ Weinberg Theory Summary: A few definitions
Bibliography
Top ~ Home
rexresearch.com
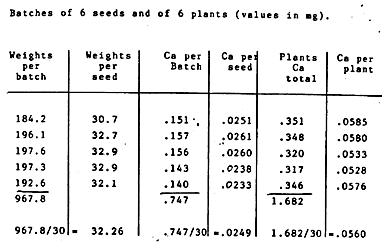{kind=link}